
Jeżeli rysunki czaszek są poprawne, to ta kwestia JEST rozwiązana. Nieważne jak daleko od siebie Scansoriopterygidae i owiraptorozaury na jakimkolwiek kladogramie kiedykolwiek wyląduję - takiego podobieństwa czaszek nie sposób tłumaczyć konwergencją.Ag.Ent pisze:Nie sądzę jednak, by kwestia pozycji filogenetycznej Scansoriopterygidae była już na dobre rozwiązana.
Archeopteryks deinonychozaurem i nomenklatura Paraves
-
d_m
Re: Archeopteryks deinonychozaurem i nomenklatura Paraves
-
Ag.Ent
- Kredowy tyranozaur

- Posty: 2349
- Rejestracja: 19 marca 2009, o 20:55
- Imię i nazwisko: Tomasz Skawiński
- Lokalizacja: Wrocław
Re: Archeopteryks deinonychozaurem i nomenklatura Paraves
Ileż to już razy twierdzono, że dany problem jest na pewno rozwiązany...
Tak wiele dinozaurów, tak mało czasu...

Re: Archeopteryks deinonychozaurem i nomenklatura Paraves
Dawno nie ospowiadałem tu, wiec post jest długi, na początek kwestie istotniejsze i szerzej omówione, dalej tylko krótkie komentarze.
Co z newsem na główną? Myślę, że z dotychczasowych postów można coś sklecić. [EDIT: ja coś zrobię]
Zob. też dalej - komentarz o ewentualnej owiraptorozaurowej naturze Scansoriopterygidae.
Nie sądzicie, że remedium na te problemy jest odrzucenie maximum parsimony i stosowanie innych metod, jak maximum likelihood?
Co myślicie o tym niżej? To post Mortimera (stąd), w którym pastwi się nad niedawną publikacją Bennetta (doi: 10.1080/08912963.2012.725727 ), najpierw cytat z Bennetta, później komentarz Mortimera:
 ), tj. wyłączenie z Deinonychosauria Troodontidae, Microraptoria, Unenlagiinae, Anchiornis+Xiaotingia, Archaeopteryx i Rahonavis przez umiejscowienie ich jako kolejno
), tj. wyłączenie z Deinonychosauria Troodontidae, Microraptoria, Unenlagiinae, Anchiornis+Xiaotingia, Archaeopteryx i Rahonavis przez umiejscowienie ich jako kolejno dalszych bliższych grup zewnętrznych wobec ptasiego kladu. Przynaleźność Scansoriopterygidae do Oviraptorosauria sugerował Paul (2010; Princeton Field Guide), Xu i in. 2010, gdzie były w Avialae (doi: 10.1007/s11434-010-4150-z ), a u Turnera i in. (2012, doi: 10.1206/748.1 ) wymuszenie pozycji Epidexipteryx wśród Oviraptorosauria wydłuża drzewo o jeden stopień.
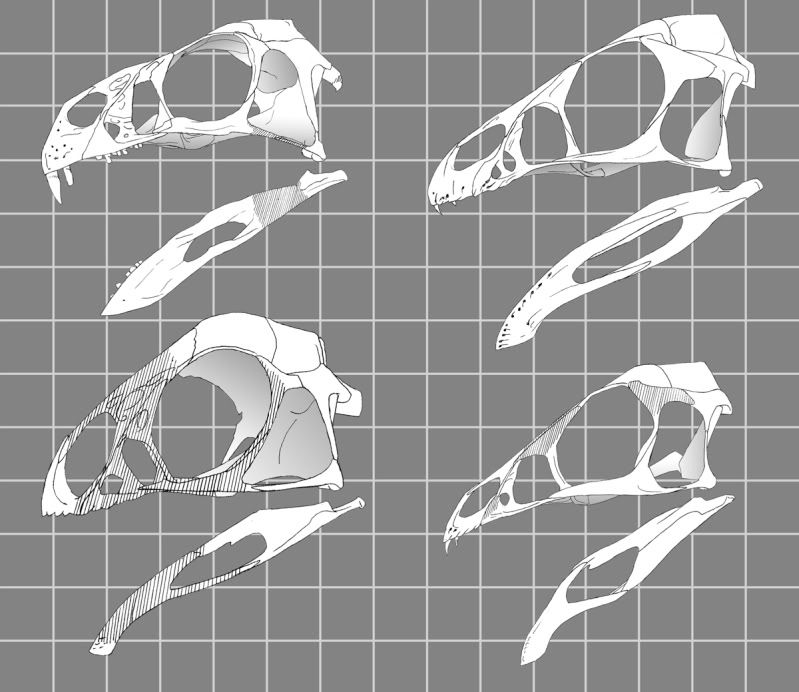
Dawid - wcześniej stwierdziłeś, że w swoim scenariuszu nie bierzesz pod uwagę Scansoriopterygidae, bo powinniśmy zakładać, że ich podobieństwo do owiraptorozaurów (i ogólnie 'morfotypu roślinożernego'), to konwergencja ;) (choć faktycznie na rysunku z Agnolina i Novasa są bardziej podobne niż w Xu i in. 2011 [Xiaotingia]).
Co by było, gdyby podobieństwo nie było tak uderzające, jak w tej rycinie (zob. też niżej inne owiraptorozaury)? Czy powinniśmy nie brać pod uwagę wielu cech, które je łączą (zob. cytowany wcześniej w wątku tekst z Xu i in. 2011)?
Similicaudipteryx jest podobny do Epidexipteryx, ale inne owiraptorozaury nie aż tak. Zob. np.
http://i16.photobucket.com/albums/b1/Qi ... odelss.jpg
(autor: Jaime Headden, qilong.wordpress.com)
Na górze Incisivosaurus i Caudipteryx, na dole Avimimus (późnokredowy i bardziej zaawansowany, reszta wczesnokredowa) i znów Caudipteryx.
Co z przedziałem czasu, który dzieli owiraptorozaury i epideksipteryksa?
Incisivosaurus jest uniwersalnie uważany za najbardziej bazalnego owiraptorozaura, później jest Caudipteryx i cała reszta. Filogeneza Similicaudipteryx jest problematyczna, został opisany jako należący do rodziny Caudipterygidae (co nie oznacza, że tworzy z nim klad) a dokładniej filogenezie tego taksonu przyjrzał się jedynie chyba Mortimer i stwierdził (w sensie zapewne tak wykazała jego niepublikowana analiza numeryczna), że to zaawansowany owiraptorozaur, zob. http://archosaur.us/theropoddatabase/Ph ... 0Taxa.html
i wpis o nim tamże. Similicaudipteryx jest młodszy (formacja Jiufotang) od Incisivosaurus, Caudipteryx i Protarchaeopteryx (formacja Yixian).
Jak już wcześniej wskazałem w tym wątku, to może być plezjomorficzna budowa Paraves+Oviraptorosauria (Xu i in. 2011, Xiaotingia).
Bardzo podobny wydaje się też Sapeornis (którego Paul włączył do Oviraptorosauria!):
A: Epidexipteryx z Agnolin i Novas
B:: Similicaudipteryx z Agnolin i Novas
C-J: Sapeornis
C: Paul 2010
D: Czerkas & Ji, 2002. A preliminary report on an omnivorous volant bird from Northeast China. Feathered Dinosaurs and the Origin of Flight. The Dinosaur Museum Journal. 1, 127-135.
E-F: Hu, Li, Hou & Xu, 2010. A new sapeornithid bird from China and its implication for early avian evolution. Acta Geologica Sinica. 84(3), 472-482.
G: Zhou & Zhang, 2003. Anatomy of the primitive bird Sapeornis chaoyangensis from the Early Cretaceous of Liaoning, China. Canadian Journal of Earth Sciences. 40, 731-747.
H: Anton Franz-Josef http://www.deviantart.com/art/Sapeornis ... -346248074
I: Hartman http://www.skeletaldrawing.com/psgallery/gallery.htm
J: ?? http://tsjok45.wordpress.com/tag/wetenschap/page/16/
Jak widać, rekonstukcje są różne, niektóre bardziej, inne mniej podobne do epidexipteryksa i similikaudipteryksa, biorąc pod uwagę faktyczny wygląd skamielin (D i E), wydaje mi się, że prawdopodobnie sapeornis był bardziej podobny do tych dwóch.
Sapeoronis jest ewidentnie ptasi i wydaje się latającym aktywnie... owiraptorozaurem?
W analizie (nie max parsimony!) macierzy Xu i in. 2011 Shenzhouraptor to takson siostrzany Epidendrosaurus+Epidexipteryx - te dwa są tam jakby 'wtrącone' do standardowej topologii bazalnych Avialae a niezgodność z czasem jest znacząca (Lee i Wothy 2012 doi: 10.1098/rsbl.2011.0884 )
Polecam zresztą dokładne porównanie całej topologii Lee i Wothyego z tą Xu i in. 2011 - ta sama macierz tylko inna metoda.
Kwestię Scansoriopterygidae w nowej pracy omawia krytycznie Headden: http://qilong.wordpress.com/2013/03/04/ ... torosaurs/
(zob. też komentarz Cau)
Mógłbyś przedstawić proponowaną przez siebie topologię?
Przygotowałem uproszczony kladogram:
legenda:
Zasięg czasowy jest tylko z grubsza.
Interesujące nas linie duchów na czerwono.
Dodałem nazwy kladów, node-based oznaczone kropą, stem-based wskazują linię (10 szczebli nazwanych, 5 nie)
Dodałem też kilka cech dotyczących rozwoju lotu, które wg Agnolina i Novasa pojawiły się w danym kladzie.
wiek Scansoriopterydigae i Achniornis jest wypośrodkowany, mogą być z batonu-kimerydu
pominąłem problematycznego Caenagnathus (?=Chirostenotes)
dodałem Haplocheirus
uwzględniłem Alnashetri jako alwarezzauryda (?wczesny cenoman)
do Microraptoria dałem Hesperonychus i Luanchuanraptor
pominąłem Wellenhoferia
Jeholornis to starszy synonim Shenzhouraptor
Dodałem informacje o geografii (grafiki z http://commons.wikimedia.org/wiki/Grey- ... tions_maps ). W ramce formy z Gondwany. Dla uproszczenia pominąłem jednak niektóre dane:
- Alvarezsauridae - formy z Ameryki Płn. i Azji są późniejsze i zaawansowane
- Microraptoria - Hesperonychus z Ameryki Płn. jest z późnej kredy
Snively & Russel (2003) "Kinematic Model of Tyrannosaurid (Dinosauria: Theropoda) Arctometatarsus Function" Journal of Morphology 255:215-227
Co z newsem na główną? Myślę, że z dotychczasowych postów można coś sklecić. [EDIT: ja coś zrobię]
No właśnie wiemy, że stopa arktometatarsalna to homoplazja z obszernych analiz filogenetycznych. Jaki jest powód, dla którego uważacie, że nie powinniśmy ciągle testować - nawet wydawałoby się odrzuconych - koncepcji? Powiedzmy, że na gruncie obecnych hipotez filogenezy stopa arktometatarsalna powstała 6 razy. Dlaczego jednak nie sprawdzić, czy przypadkiem 4? Ta cecha, zapewne przy wsparciu innych, może wszak spowodować, że pewne taksony się do siebie zbliżą, czego byśmy nie zaobserwowali, odrzucając tą cechę z góry. Wydawałoby się, że stopa arktometatarsalna to cecha, która nie powstaje w ewolucji łatwo...Utahraptor pisze:Skoro wiemy, że arktometatarsalna stopa powstała wiele razy w ewolucji, to poco ją stosować, gdy porównujemy ze sobą celurozaury? Żeby zbliżała nam sztucznie ornitomimozaury i tyranozaury? Poco stosować homoplazje w analizach? Jest to zaprzeczenie kladystyki.nazuul pisze:To co byś proponował jako rozwiązanie tego problemu?Utahraptor pisze:Dlatego część osób całkowicie odrzuca tę metodę. Ja nie jestem aż tak radykalny
Ja skłaniam się do stosowania mniejszej ilości cech, które nie powstają w ewolucji łatwo i nad którymi można się spokojnie zastanowić. Używając ogromnych macierzy nie jesteśmy w stanie zastanawiać się nad stosowanymi cechami. Wyrzuciłbym z macierzy cechy ewidentnie homoplastyczne.
Zob. też dalej - komentarz o ewentualnej owiraptorozaurowej naturze Scansoriopterygidae.
Nie sądzicie, że remedium na te problemy jest odrzucenie maximum parsimony i stosowanie innych metod, jak maximum likelihood?
Co myślicie o tym niżej? To post Mortimera (stąd), w którym pastwi się nad niedawną publikacją Bennetta (doi: 10.1080/08912963.2012.725727 ), najpierw cytat z Bennetta, później komentarz Mortimera:
Mortimer pisze:I wholeheartedly agree that we need to spend more time formulating and testing characters, but find the rest laughable. First, there have been numerous papers showing adding taxa and characters usually results in a more accurate tree, though there are eventual limits to the payoff for both. Bennett's 19 taxa and 144 characters is nowhere near those limits, needless to say. What's truly absurd is that Bennett thinks he knows the angle of the anteromedial astragalar corner is less likely to change than the presence of an antorbital fenestra. How would you even determine that? Homoplasy? Well, the corner character never converges or reverses in Nesbitt's matrix, so that can't be the reason. The number of mutations it takes? We have no clue about that, and given recent studies showing single genes can have huge phenotypic effects, I think it would be foolish to equate the apparent significance of a character with its phylogenetic worth. The same issue surrounds his choice to make some characters irreversable. How does he know which reversals are impossible genetically? What Bennett is advocating is a return to the good ol' days of "key characters", or in this case key character complexes, which are even worse.Bennett, 2012 pisze:The general trend in phylogenetic analyses of diapsids has been towards analyses with more taxa and more
characters, as if more necessarily equated to better, and Nesbitt’s (2011) analysis is but the most recent. Unfortunately, the usual way to get more characters is to atomise the morphology of structures, coding for more and more insignificant features of structures, which leads to multiple unintentional weightings of the underlying character. Nesbitt’s analysis has 30 characters from the astragalus and calcaneum including, for example the shape of the anteromedial corner of the astragalus: acute or obtuse. It is absurd that the angle of the corner, a few degrees this way or that has the same weight in an analysis as the presence of an antorbital fenestra, a calcaneal tuber or a pteroid bone. I reject such atomisation of morphology, and point out again that most effort should go into formulation and testing of characters, rather than the analysis of the data and manipulation of trees and that one should use characters only if the ‘hypotheses of homology cannot be refuted’ (Gaffney et al. 1991).
Kwestią wstępu, to osobiście uważam, że najistotniejsza jest 'masowa parafiletyzacja' (Dawid Mazurek pisze:Jeżeli rysunki czaszek są poprawne, to ta kwestia JEST rozwiązana. Nieważne jak daleko od siebie Scansoriopterygidae i owiraptorozaury na jakimkolwiek kladogramie kiedykolwiek wyląduję - takiego podobieństwa czaszek nie sposób tłumaczyć konwergencją.Ag.Ent pisze:Nie sądzę jednak, by kwestia pozycji filogenetycznej Scansoriopterygidae była już na dobre rozwiązana.
 ), tj. wyłączenie z Deinonychosauria Troodontidae, Microraptoria, Unenlagiinae, Anchiornis+Xiaotingia, Archaeopteryx i Rahonavis przez umiejscowienie ich jako kolejno
), tj. wyłączenie z Deinonychosauria Troodontidae, Microraptoria, Unenlagiinae, Anchiornis+Xiaotingia, Archaeopteryx i Rahonavis przez umiejscowienie ich jako kolejno Dawid - wcześniej stwierdziłeś, że w swoim scenariuszu nie bierzesz pod uwagę Scansoriopterygidae, bo powinniśmy zakładać, że ich podobieństwo do owiraptorozaurów (i ogólnie 'morfotypu roślinożernego'), to konwergencja ;) (choć faktycznie na rysunku z Agnolina i Novasa są bardziej podobne niż w Xu i in. 2011 [Xiaotingia]).
Co by było, gdyby podobieństwo nie było tak uderzające, jak w tej rycinie (zob. też niżej inne owiraptorozaury)? Czy powinniśmy nie brać pod uwagę wielu cech, które je łączą (zob. cytowany wcześniej w wątku tekst z Xu i in. 2011)?
Similicaudipteryx jest podobny do Epidexipteryx, ale inne owiraptorozaury nie aż tak. Zob. np.
http://i16.photobucket.com/albums/b1/Qi ... odelss.jpg
(autor: Jaime Headden, qilong.wordpress.com)
Na górze Incisivosaurus i Caudipteryx, na dole Avimimus (późnokredowy i bardziej zaawansowany, reszta wczesnokredowa) i znów Caudipteryx.
Co z przedziałem czasu, który dzieli owiraptorozaury i epideksipteryksa?
Incisivosaurus jest uniwersalnie uważany za najbardziej bazalnego owiraptorozaura, później jest Caudipteryx i cała reszta. Filogeneza Similicaudipteryx jest problematyczna, został opisany jako należący do rodziny Caudipterygidae (co nie oznacza, że tworzy z nim klad) a dokładniej filogenezie tego taksonu przyjrzał się jedynie chyba Mortimer i stwierdził (w sensie zapewne tak wykazała jego niepublikowana analiza numeryczna), że to zaawansowany owiraptorozaur, zob. http://archosaur.us/theropoddatabase/Ph ... 0Taxa.html
i wpis o nim tamże. Similicaudipteryx jest młodszy (formacja Jiufotang) od Incisivosaurus, Caudipteryx i Protarchaeopteryx (formacja Yixian).
Jak już wcześniej wskazałem w tym wątku, to może być plezjomorficzna budowa Paraves+Oviraptorosauria (Xu i in. 2011, Xiaotingia).
Bardzo podobny wydaje się też Sapeornis (którego Paul włączył do Oviraptorosauria!):
B:: Similicaudipteryx z Agnolin i Novas
C-J: Sapeornis
C: Paul 2010
D: Czerkas & Ji, 2002. A preliminary report on an omnivorous volant bird from Northeast China. Feathered Dinosaurs and the Origin of Flight. The Dinosaur Museum Journal. 1, 127-135.
E-F: Hu, Li, Hou & Xu, 2010. A new sapeornithid bird from China and its implication for early avian evolution. Acta Geologica Sinica. 84(3), 472-482.
G: Zhou & Zhang, 2003. Anatomy of the primitive bird Sapeornis chaoyangensis from the Early Cretaceous of Liaoning, China. Canadian Journal of Earth Sciences. 40, 731-747.
H: Anton Franz-Josef http://www.deviantart.com/art/Sapeornis ... -346248074
I: Hartman http://www.skeletaldrawing.com/psgallery/gallery.htm
J: ?? http://tsjok45.wordpress.com/tag/wetenschap/page/16/
Jak widać, rekonstukcje są różne, niektóre bardziej, inne mniej podobne do epidexipteryksa i similikaudipteryksa, biorąc pod uwagę faktyczny wygląd skamielin (D i E), wydaje mi się, że prawdopodobnie sapeornis był bardziej podobny do tych dwóch.
Sapeoronis jest ewidentnie ptasi i wydaje się latającym aktywnie... owiraptorozaurem?
W analizie (nie max parsimony!) macierzy Xu i in. 2011 Shenzhouraptor to takson siostrzany Epidendrosaurus+Epidexipteryx - te dwa są tam jakby 'wtrącone' do standardowej topologii bazalnych Avialae a niezgodność z czasem jest znacząca (Lee i Wothy 2012 doi: 10.1098/rsbl.2011.0884 )
Polecam zresztą dokładne porównanie całej topologii Lee i Wothyego z tą Xu i in. 2011 - ta sama macierz tylko inna metoda.
Kwestię Scansoriopterygidae w nowej pracy omawia krytycznie Headden: http://qilong.wordpress.com/2013/03/04/ ... torosaurs/
(zob. też komentarz Cau)
Też mam czasem wrażenie, że kladogramy są na odwrót.Dawid Mazurek pisze:Podobają mi się pewne aspekty przedstawionej topologii, ale jeśli weźmie się pod uwagę czas geologiczny, moim zdaniem linia ewolucyjna wiodąca do ptaków jest zakorzenione do góry nogami i w rzeczywistości analizowany zapis kopalny nie dokumentuje wznoszenia się w powietrze, tylko kolejne etapy schodzenia na ziemię połączone z utratą kolejnych ptasich synapomorfii.
Mógłbyś przedstawić proponowaną przez siebie topologię?
Przygotowałem uproszczony kladogram:
Zasięg czasowy jest tylko z grubsza.
Interesujące nas linie duchów na czerwono.
Dodałem nazwy kladów, node-based oznaczone kropą, stem-based wskazują linię (10 szczebli nazwanych, 5 nie)
Dodałem też kilka cech dotyczących rozwoju lotu, które wg Agnolina i Novasa pojawiły się w danym kladzie.
wiek Scansoriopterydigae i Achniornis jest wypośrodkowany, mogą być z batonu-kimerydu
pominąłem problematycznego Caenagnathus (?=Chirostenotes)
dodałem Haplocheirus
uwzględniłem Alnashetri jako alwarezzauryda (?wczesny cenoman)
do Microraptoria dałem Hesperonychus i Luanchuanraptor
pominąłem Wellenhoferia
Jeholornis to starszy synonim Shenzhouraptor
Dodałem informacje o geografii (grafiki z http://commons.wikimedia.org/wiki/Grey- ... tions_maps ). W ramce formy z Gondwany. Dla uproszczenia pominąłem jednak niektóre dane:
- Alvarezsauridae - formy z Ameryki Płn. i Azji są późniejsze i zaawansowane
- Microraptoria - Hesperonychus z Ameryki Płn. jest z późnej kredy
Rozumiem, że chodzi o to, że jestem zwolennikiem nomenklatury filogenetyczej i metody badania filogenezy za pomocą numerycznych analiz filogenetyczych. Nie wydaje mi się, żeby to była kwestia przyjęcia przeze mnie aktualnej mody... Mógłbyś podać konkretne przykłady, które potwierdzają Twoje słowa?Dawid Mazurek pisze:Mam wrażenie, że często w dyskusjach brakuje Ci szerszego spojrzenia - chodzi mi o "naukowe mody", paradygmaty i ewolucję podejść; przykładowo - trzeba uwzględniać najnowsze propozycje filogenetyczne i nomenklatoryczne, ale nie ma się co do dzisiejszego podejścia do systematyki przyzwyczajać jako do paradygmatu ostatniego.
Dlaczego wczesna jura czy trias?Utahraptor pisze:Początki maniraptorów sięgają na pewno środkowej lub wczesnej juryAg.Ent pisze:Jeśli stosować definicje Agnolina i Novasa, to na pewno nie ;) Zresztą w ogóle przyjęta przez nich nomenklatura, a zwłaszcza zmienianie powszechnie przyjętej, jest bez sensu.Dawid Mazurek pisze:Początki Avialae sięgają wczesnej jury lub triasu.
Niezbyt wiadomo, które ze znanych form to bazalne deinonychozaury i czy w ogóle jakieś znamyUtahraptor pisze:Bazalne dromeozaurydy być może, ale nie bazalne deinonychozaurynazuul pisze:z biegiem to różnie bywało. kierując się proporcjami dłogości kości i formą śrdóstopia, to zaawansowane dromeozaurydy (Eudromaeosauria) i owiraptorozaury (Oviraptoridae) były gorzej biegające niż te bardziej bazalne, u alwarezauroidów i troodontydów było rozwinięcie szybkiego biegu.Utahraptor pisze:podejrzewam, że deinonychozaury, alwarezzaurydy i owiraptorozaury reprezentują kolejne etapy schodzenia z drzew. Świadczą o tym liczne paralelizmy ewolucyjne, będące pewnie skutkiem wspólnych preadaptacji do lotu oraz pióra. Zdaniem dr Gerarda Gierlińskiego tropy owiraptorozaurów są niemal identyczne z ptasimi. Jeśli mam rację, to formy bazalne powinny słabiej biegać, natomiast formy bardziej zaawansowane lepiej. Gdyby działał WAIR tego trendu by nie było.
Jednak arctometatarsus to element ruchomy, zob. np.Utahraptor pisze:Śródstopie jest u dzisiejszych ptaków zrośnięte. Subarctometatarsus jest dla mnie po prostu stanem pośrednim, gdzie kości śródstopia są gęsto upakowane. Jeżeli jest to synapomorfia owiraptorozaurów i Paraves i jest adaptacją do lądowania, to potwierdza to hipotezę, że to wszystko są potomkowie nadrzewnych archozaurów (teropodów?).nazuul pisze:Dlaczego tak sądzisz?Utahraptor pisze:U mikroraptora jest to [arktometatarsalna stopa] jednak nie w pełni rozwinięte i może być efektem tendencji do zrastania się śródstopia we wczesnej ewolucji ptaków.
Snively & Russel (2003) "Kinematic Model of Tyrannosaurid (Dinosauria: Theropoda) Arctometatarsus Function" Journal of Morphology 255:215-227
Moim zdaniem powinno się coś zmieniać tylko wtedy, gdy jest to szczególnie uzasadnione (o ile oczywiście definicje są dobre), np. w razie drastycznego rozszerzenia liczby taksonów, które należą do danej, w określony sposób zdefiniowanej, nazwy kladu (np. nie jest dla mnie problemem, że skansoriopterygidy należą do Oviraptorosauria, ale gdyby np. miało być odwrotnie, tj. Oviraptorosauria należą do Scansoriopterygidae, to definicje do zmiany).Ag.Ent pisze:Z "kuciem żelaza póki gorące" trudno mi się zgodzić - drobne modyfikowanie definicji nie jest niczym złym, ale nie powinno się "zaprzeczać" koncepcji, którą miał dany autor, kiedy tworzył daną nazwę (niestety zdarzały się takie przypadki).Dawid Mazurek pisze:Co do uniwersalności nazw i zmienianiu ich znaczenia. Wiem o co Ci chodzi i pewnie w tym przypadku były to błędy autorów, ale generalnie klady dinozaurów to nie są jakieś rozpoznawalne od stuleci wydzielenia, ich podział dopiero się wykluwa (większość to osiągnięcia ostatnich kilkudziesięciu, czy nawet kilku lat), więc czemu by ich nie modyfikować, kuć żelazo póki gorące?
określony zestaw taksonów = specyfikatoryDawid Mazurek pisze:Poza tym, jak już gdzieś wcześniej pisałem, bardziej pożądana stabilność to ta, by nazwa była przywiązana do określonego zestawu taksonów niż do określonego przez specyfikatory wycinku drzewa.
Faktycznie wysyp nowych nazw i często grzebanie w definicjach jest zupełnie niepotrzebne. Myślę że w ostatnich kilku latach w nieptasiodinozaurologii jest tylko kilka sensownych propozycji nowych nazw, reszta jest niepotrzebna.Utahraptor pisze:Jeżeli chodzi o nomenklaturę, to się nie znam, ale tworzenie nowych nazw kladów rzeczywiście wydaje się zbędne. Z Dawidem ostrzegaliśmy, że nazwy kladów będą się mnożyć.

{kind=link}
-
d_m
Re: Archeopteryks deinonychozaurem i nomenklatura Paraves
Tyle nowych danych - jak przerobię, to się wypowiem Kolejny raz okazuje się, że nic nie jest takie proste jak mogłoby się na początku wydawać... ;-)
Re: Archeopteryks deinonychozaurem i nomenklatura Paraves
Z dotychczasowych wypowiedzi w wątku skleciłem informację na główną, w wersji konsensusowej, dodałem trochę - zwracam uwagę zwłaszcza na notkę o Haplocheirus. Jeśli należy coś zmienić, zwłaszcza w 'konsensusowości' tekstu, to koniecznie piszcie, najlepiej konkretne propozycje zmian (obecny tekst->proponowany tekst). Podrubione to sugerowane odesłania do ED, nie wszystkie z tych haseł istnieją (paralelizm, symplezjomorfia - te dwa wg MT są, nie ma homoplazja + kilka rodzajów), wiec nie wiem czy je linkować - dałem je z podkreśleniem. W nawiasach kwadratowych dałem hasło jeśli nie jest oczywiste, do którego ma linkować dane słowo. Autorzy: MZ, DM, MT, TS, siebie dałem na początek z uwagi na zrobienie obrazka i jego podpisu oraz opracowanie tekstu (można też dać sapeornisa), DM i MT alfabetycznie:
MZ, DM, MT, TS pisze:Ukazało się od dawna oczekiwane badanie dwójki argentyńskich paleontologów - Federico Agnolína i Fernando Novasa, dotyczące wczesnej ewolucji ptaków. Prezentujemy nasze opinie dotyczące tej publikacji, wyrażone podczas dyskusji na forum.
Wiele cech uznawanych dotychczas za synapomorfie danych kladów, autorzy uważają za o wiele szerzej rozprzestrzenione - powstałe wcześniej lub nabyte konwergentnie. Podoba nam się nam to, że analiza filogenetyczna została tu użyta do przedstawienia scenariusza ewolucyjnego z propozycją przemian anatomicznych. Istotne wydają się szczególnie dwa aspekty przedstawionej hipotezy.
Pierwszy to włączenie jurajskich skansoriopterygidów: Epidexipteryx i Epidendrosaurus do owiraptorozaurów (nie jest to całkiem niespodziewane, już wcześniej poważnie rozpatrywano tę opcję, ale wydawało się, że lepiej wspierana jest jednak bardziej "ptasia" wersja). Jeśli Scansoriopterygidae są owiraptorozaurami, to znika spora luka w zapisie kopalnym.
Drugi istotny aspekt hipotezy Agnolína i Novasa to wyłączenie z Deinonychosauria Troodontidae, Microraptoria, Unenlagiinae, Anchiornis+Xiaotingia, Archaeopteryx i Rahonavis przez umiejscowienie ich jako kolejno bliższych krewniaków ptaków ('masowa parafiletyzacja [?? grupa parafiletyczna]' Deinonychosauria) (zob. rysunek). Jako bardziej zaawansowane od Dromaeosauridae i Troodontidae, lecz problematyczne, autorzy uznali też niektóre dotychczasowe deinonychozaury: Shanag, Mahakala, Hulsanpes, Tianyuraptor, Jinfengopteryx, Pamparaptor oraz Luanchuanraptor, który ma być ostatnim znanym przedstawicielem Microraptoria. Także problematycznego Unquillosaurus włączono do Avialae.
Wg części z nas, kształt drzewa i cechy, które za nią stoją są jednak zupełnie chybione. Pokazuje to, jak mało warte są kladogramy oparte na macierzach zawierających jakieś przypadkowe proporcje i kształty kości kończyn i czaszki. Autorzy zaserwowali nam zupę synapomorfii, paralelizmów, symplezjomorfii i homoplazji.
Możliwe, że zaawansowana budowa troodontydów i dromeozaurydów została zinterpretowana jako prymitywna, a plezjomorficzna [plezjomorfia] budowa aweraptorów jako zaawansowana (Averaptora to nazwa zaproponowana przez autorów dla identycznego kladu, co Avialae w swojej pierwotnej i chyba najpopularniejszej koncepcji, tj. jako zwierzęta bliższe współczesnym ptakom niż dromeozaurydom). W rezultacie mamy schemat, który zupełnie nie ima się biogeografii, co autorzy przyznają pod koniec rozdziału. Podobnie jest z czasem geologicznym - proponowana kolejność odgałęzień ma się nijak do zapisu kopalnego. Jeśli weźmie się pod uwagę czas geologiczny, to linia ewolucyjna wiodąca do ptaków może być zakorzeniona 'do góry nogami' i w rzeczywistości analizowany zapis kopalny nie dokumentuje wznoszenia się w powietrze, tylko kolejne etapy schodzenia na ziemię połączone z utratą kolejnych ptasich synapomorfii. Możliwe, że maniraptory były pierwotnie nadrzewnymi zwierzętami. Być może zeszły na ziemię kilka razy, dając kilka osobnych grup - owiraptorozaurów, alwarezzauroidów, deinonychozaurów. Szczególnie istotne wydaje się tutaj włączenie Scansoriopterygidae do Oviraptorosauria. Sugerowana wcześniej nadrzewność Scansoriopterygidae została ostatnio wzmocniona nowymi danymi, do których Agnolín i Novas nie mieli dostępu. U Scansoriopteryx (=Epidendrosaurus) zaobserwowano wydłużone pióra na śródręczu i śródstopiu, co sugeruje, że on także był czteroskrzydłym lotnikiem (Stephen Czerkas w: Feduccia, A. 2013. Bird origins anew. The Auk 130 (1): 1-12). Zatem pogląd autorów, że lot powstał raz, lecz stopniowo, może być nietrafny. Nie można też wykluczyć, że hipoteza owiraptorozaurowa zostanie potwierdzona, a Scansoriopterygidae okażą się np. monofiletyczną grupą [klad] wewnątrz Oviraptorosauria, która weszła na drzewa, podczas gdy pozostałe owiraptorozaury "poszły" w kierunku kursorialności, odziedziczając pewne preadaptacje ku nadrzewności.
Trzeba zauważyć, że w analizie zabrakło kilku istotnych taksonów, np. Haplocheirus, który jest wczesnym, ale typowo lądowym przedstawicielem Alvarezsauroidea. Alwarezzauroidy w hipotezie Agnolína i Novasa znajdują się pomiędzy owiraptorozaurami a kladem (Troodontidae(Dromaeosauridae+Avialae)), co zaburza przedstawiony wyżej obraz kolejnego schodzenia z drzew kilku grup. Pozycja Alvarezsauroidea w filogenezie jest jednak niepewna a sam Haplocheirus może być przedstawicielem bardziej bazalnej grupy - Ornithomimosauria.
Wg części z nas, kwestia pozycji filogenetycznej Scansoriopterygidae nie jest jeszcze na dobre rozwiązana. Autorzy wskazali, jak bardzo podobne są czaszki jurajskiego skansoriopterygida Epidexipteryx i wczesnokredowego owiraptorozaura Similicaudipteryx. Sugeruje to, że - jeśli ryciny oddają rzeczywistą budowę - te formy muszą być blisko spokrewnione. Jednak inne wczesnokredowe owiraptorozaury nie są aż tak podobne (Caudipteryx czy Incisivosaurus, który jest uniwersalnie uważany za najbardziej bazalnego owiraptorozaura). Możliwe też, że taka budowa jest plezjomorficzna dla kladu Paraves+Oviraptorosauria (=Chuniaoae). Bardzo podobny wydaje się też wczesokredowy ptak Sapeornis (którego Gregory Paul w swojej popularnonaukowej książce "The Princeton Field Guide To Dinosaurs" włączył nawet do Oviraptorosauria!). Sapeornis jest bardzo ptasi i wydaje się latającym aktywnie... no właśnie - ptakiem? owiraptorozaurem?
Podsumowując, zaprezentowana koncepcja filogenetyczna jest fajna, bo czuć powiew świeżości. Jednak zapewne jeszcze długo dokładny przebieg wczesnej ewolucji ptaków (na granicy z 'klasycznymi' dinozaurami) nie będzie mógł być uważany za ostatecznie rozwiązany.
Zmodyfikowany opis proponowanego obrazka:
http://www.forum.dinozaury.com/download ... &mode=view
Filogeneza Maniraptora wg Agnolína i Novasa (2013). Zaznaczono: nazwy kladów - z pominięciem uznanych przez nas za niepotrzebne zmian Agnolína i Novasa w definicjach filogenetycznych (node-based oznaczone kropą, stem-based wskazują linię); kilka cech dotyczących rozwoju lotu, które wg Agnolína i Novasa pojawiły się w danym kladzie; informacje o geografii (w ramce formy z Gondwany; dla uproszczenia pominięto niektóre taksony: w Alvarezsauridae - formy z Ameryki Płn. i Azji, gdyż są późniejsze i zaawansowane oraz w Microraptoria Hesperonychus z Ameryki Płn., gdyż jest z późnej kredy); istotne luki w zapisie kopalnym zaznaczono na czerwono. 'Tetrapteryx' oznacza powstanie morfologii 'czteroszkszydłego lotnika', ze znakiem zapytania za Czerkas w Feduccia, 2013. Pominęto problematyczne: Caenagnathus (?=Chirostenotes), Wellnhoferia (?=Archaeopteryx). Dodano Haplocheirus i uwzględniono Alnashetri jako alwarezzauryda (wczesny cenoman?) oraz Hesperonychus i Luanchuanraptor jako mikroraptory. Nie dodano 'Lori' - nienazwanego troodontyda z późnej jury Ameryki Płn. opisanego w abstrakcie z konferencji SVP z 2005 r. (jego filogeneza jest niepewna biorąc pod uwagę m.in. wyłączenie z Troodontidae Jinfengopteryx). Wiek Scansoriopterygidae i Anchiornis został wyśrodkowany (mogą być z batonu-kimerydu). Tabela stratygraficzna z International Chronostratigraphic Chart, International Commission on Stratigraphy, 2012 (grafika przygotowana przez K.M. Cohen, S. Finney, P.L. Gibbard); grafiki kontynentów z http://commons.wikimedia.org/wiki/Grey- ... tions_maps
-
d_m
Re: Archeopteryks deinonychozaurem i nomenklatura Paraves
Testowanie tego, czy stopa arktometatarsalna powstała raz czy wiele razy to jedna sprawa (osobny artykuł), a filogeneza teropodów to druga. Jeżeli konsens będzie taki, że powstała raz, można jej będzie używać w pracach o filogenezie, a jak odmienny - cecha wylatuje. Nie kodujemy obecności piór i włosów jako tego samego stanu tej samej cechy, ale jeśli ktoś wykaże ich homologię, może będziemy musieli zacząć...Jaki jest powód, dla którego uważacie, że nie powinniśmy ciągle testować - nawet wydawałoby się odrzuconych - koncepcji?
Można zrobić nakładając tę cechę na już zrobiony bez użycia homoplazji kladogram.Powiedzmy, że na gruncie obecnych hipotez filogenezy stopa arktometatarsalna powstała 6 razy. Dlaczego jednak nie sprawdzić, czy przypadkiem 4?
Dokładnie. Komputerowa analiza filogenezy ma charakter statystyczny, probabilistyczny, więc używanie śmieciowych danych wejściowych może powodować sztuczne zbliżanie/oddalanie się taksonów, czego byśmy nie zaobserwowali używając "dobrych" cech.Ta cecha, zapewne przy wsparciu innych, może wszak spowodować, że pewne taksony się do siebie zbliżą, czego byśmy nie zaobserwowali, odrzucając tą cechę z góry.
Dokładnie. Cechy nie są równowartościoweWydawałoby się, że stopa arktometatarsalna to cecha, która nie powstaje w ewolucji łatwo...
Co ktoś z nas może sądzić, a jak się w praktyce tworzy macierze i przeprowadza analizy... dwie różne sprawy.Nie sądzicie, że remedium na te problemy jest odrzucenie maximum parsimony i stosowanie innych metod, jak maximum likelihood?
Podoba mi się co napisał Bennett i generalnie się pod tym podpisuję, a omawiany przez nas przypadek jaszczurek stoi w sprzeczności z tymi słowami Mortimera: "First, there have been numerous papers showing adding taxa and characters usually results in a more accurate tree". Ja bym powiedział, że dodawanie pojedynczych cech i taksonów do już istniejących macierzy (lub po prostu podobny schemat budowania nowych macierzy) z natury rzeczy zazwyczaj nie ma wystarczającej siły statystycznej by obrócić topologię o 180 stopni, więc podrasowana analiza jest zgodna w ogółach ze swoją poprzedniczką.
Nie pamiętam co napisałem, ale bardziej mi chodziło o to, że póki nie za wiele wiemy o tym co to za grupa, jaka jest morfologia, pozycja systematyczna, rozprzestrzenienie czasowe i geograficzne, dla przejrzystości wywodu można je pominąć, tak jak czasem z macierzy wycina się niekompletne taksony, które destabilizują topologię... Alternatywnie: JA niewiele o niej wiem, więc ją pomijam, upraszczam, myślę syntetycznie...Dawid - wcześniej stwierdziłeś, że w swoim scenariuszu nie bierzesz pod uwagę Scansoriopterygidae, bo powinniśmy zakładać, że ich podobieństwo do owiraptorozaurów (i ogólnie 'morfotypu roślinożernego'), to konwergencja
Dopiero co był wyjazd do Hamburga, więc ciężko będzie skołować kolejną kasę na wyjazd w perspektywiczny region o pożądanym wieku, ale pomysł dobryCo z przedziałem czasu, który dzieli owiraptorozaury i epideksipteryksa?
No skoro tak, to faktycznie dużo to zmienia, a więc pozycja w obrębie owiraptorozaurów, lub informatywna dla badania początków większego kladu (np. Paraves+Oviraptorosauria) plezjomorficzna budowa, być może ujawniona w związku z wiekiem ontogenetycznym, kto wie, może wręcz 2/3 Scansoriopterygidae to po prostu polifiletyczna grupa pisklaków? To by pasowało!!! Wiemy też, że wszystkie 3 okazy mogą być synonimiczne - dwa pisklaki i jeden dorosły o cechach pisklaków/przodków. Tak tylko głośno myślę... Innych możliwości nie widzę, no chyba że rekonstrukcje czaszek są tendencyjne...Jak już wcześniej wskazałem w tym wątku, to może być plezjomorficzna budowa Paraves+Oviraptorosauria (Xu i in. 2011, Xiaotingia).
Bardzo podobny wydaje się też Sapeornis (którego Paul włączył do Oviraptorosauria!)
Innych rewelacji kladogramowych (e.g. Sapeornis) nie skomentuję, bo za mało wiem o anatomii tych form.
Co do komentarza Headena: jestem pod wrażeniem ile pracy włożył w rozpracowanie każdej z komentowanych cech na czynniki pierwsze. Nie znam się na osteologii, więc nie będę się wczytywał i roboczo uznam, że ma rację
Poczytam coś więcej o Sapeornis
Nie chce mi się bazgrać, więc tak w skrajnym uproszczeniu: późna jura - najstarsze znane ptaki (linia Archaeopteryx i Anchiornis) współwystępują z pierwszymi wtórnymi nielotami (Scansoriopterygidae i ich owiraptorozaurzy potomkowie). Wczesna kreda - druga fala wtórnych nielotów (tetrapteryksy) i dalej z nich troodony, z których z kolei w później kredzie dromeozaury. Gdyby taki scenariusz był prawdziwy (a wątpię, aż tak prosto nie ma), to stosując definicje musielibyśmy powiedzieć, że troodony i dromeozaury powstały gdzieś w środkowej kredzie, a wcześniejsze 'troodony' to w rzeczywistości bazalne deinonychozaury. Ja wolę jednak by taksonomia nie pomijała aspektów anatomicznych i powiedziałbym że przy takim scenariuszu dromeozaury wydzieliły się z troodonów, które trwały nadal. Terizinozaury i alwarezzaurydy? Nie wiem, nie mam pomysłu na tą chwilę, musiałbym polookać więcej w drzewka...Też mam czasem wrażenie, że kladogramy są na odwrót.
Mógłbyś przedstawić proponowaną przez siebie topologię?
Nie tylko o to mi chodzi. Zarzuciłeś któremuś z moich pomysłów, że nie bierze pod uwagę Scansoriopterygidae. Ja w takim momencie mówię: poczekajmy, będzie 10 gatunków i konsens co do ich pozycji filogenetycznej, weźmie się je pod uwagę. Nie ma co się trzymać ustaleń sprzed roku, bo co kladogram trzeba by zmieniać scenariusz.Rozumiem, że chodzi o to, że jestem zwolennikiem nomenklatury filogenetyczej i metody badania filogenezy za pomocą numerycznych analiz filogenetyczych. Nie wydaje mi się, żeby to była kwestia przyjęcia przeze mnie aktualnej mody... Mógłbyś podać konkretne przykłady, które potwierdzają Twoje słowa?
Daję ewolucji czas na zrobienie archeopteryksa, biorę pod uwagę prawdopodobną ghost lineage (szybujący przodek archeopteryksa z jakichś lasów na archipelagu wysp z którego nic w zapisie kopalnym nie przetrwało; pomyśl o rozprzestrzenieniu geograficznym dzisiejszych organizmów z perspektywami na aktywnie latających potomków), no i problematyczne jeżeli chodzi o datowanie ptasie (lub nie ptasie) tropy.Dlaczego wczesna jura czy trias?
No gdyby tak było, że specyfikatorami w przypadku archozaurów byłyby i jakiś fitozaur i jakiś aetozaur i jakiś dinozaur i... to OKokreślony zestaw taksonów = specyfikatory
Jak topologie drzew się ustabilizują, to znajdą się ludzie, którzy zaczną nazywać każde odgałęzienie, powstanie kupa nowych nazw, których nikt nie spamięta i po samej nazwie nie będzie w stanie powiedzieć, czy to jakaś ważna grupa, stąd jestem za tworzeniem jednostek parafiletycznych i nadawanie im rang. W przypadku trzech kolejnych taksonów leżących coraz bardziej w stronę korony (A(B(C(D-Z)))), lepiej utworzyć dla nich jedną jednostkę o randze mówiącej że to SUBIEKTYWNIE mała grupa, czyli zrobić rodzinę dla A, B i C, niż nazywać każdy z kladów z osobna.Myślę że w ostatnich kilku latach w nieptasiodinozaurologii jest tylko kilka sensownych propozycji nowych nazw, reszta jest niepotrzebna.
EDIT: bardzo fajny news na główną, nazuul
Re: Archeopteryks deinonychozaurem i nomenklatura Paraves
Odpowiem później, póki co chciałbym się dowiedzieć, czy są jakies uwagi do proponowanego newsa (i tak już spóźnionego)?
-
d_m
Re: Archeopteryks deinonychozaurem i nomenklatura Paraves
Moim zdaniem jest OK, lepszego konsensu nie osiągniemy ;P
-
Ag.Ent
- Kredowy tyranozaur
- Posty: 2349
- Rejestracja: 19 marca 2009, o 20:55
- Imię i nazwisko: Tomasz Skawiński
- Lokalizacja: Wrocław
Re: Archeopteryks deinonychozaurem i nomenklatura Paraves
Moim zdaniem też ciężko będzie o coś lepszego (choć nie ze wszystkim się zgadzam, ale to normalne w dyskusji ;)). Dobra robota!
Kilka literówek tylko:
Kilka literówek tylko:
Alvarezsauroidea.Pozycja Alvarezsaroidea w filogenezie jest jednak niepewna a sam Haplocheirus może być przedstawicielem bardziej bazalnej grupy - Ornithomimosauria.
SapeornisSapeoronis jest bardzo ptasi i wydaje się latającym aktywnie... no właśnie - ptakiem? owiraptorozaurem?
WellnhoferiaWellenhoferia (?=Archaeopteryx)
definicjachnazwy kladów - z pominięciem uznanych przez nas za niepotrzebne zmian Agnolína i Novasa w deficjach filogenetycznych
Scansoriopterygidae, AnchiornisWiek Scansoriopterydigae i Achniornis został wyśrodkowany
preadaptacje (albo egzaptacje)pozostałe owiraptorozaury "poszły" w kierunku kursorialności, odziedziczając pewne preaptacje ku nadrzewności.
-
Utahraptor
- Moderator

- Posty: 2600
- Rejestracja: 22 października 2007, o 18:29
Re: Archeopteryks deinonychozaurem i nomenklatura Paraves
Tekst fajny i kompromisowy ;)
Boję się tylko, że jest najeżony zbyt specjalistycznym słownictwem, ale i tak się starałeś wszystko wyjaśniać. Paralelizmy i symplezjomorfie chyba opisałem w ED.
Do dyskusji również niedługo wrócę ;)
Boję się tylko, że jest najeżony zbyt specjalistycznym słownictwem, ale i tak się starałeś wszystko wyjaśniać. Paralelizmy i symplezjomorfie chyba opisałem w ED.
Do dyskusji również niedługo wrócę ;)
Biologia, UW
Re: Archeopteryks deinonychozaurem i nomenklatura Paraves
to umówmy się tak, że jeśli ich nie ma, to je niezwłocznie napiszesz, kiedy ED wróci (+jeszcze najlepiej homoplazja)Utahraptor pisze:Paralelizmy i symplezjomorfie chyba opisałem w ED.
Natomiast ja wrzucę zaktualizowane Shanag i Luanchuanraptor (Hulsanpes jednak jest w ED; hasła o grupach teropodów też oczywiście zaktualizuję szybko).
Zaś Tomek wrzuci Similicaudipteryx, Dawid Mika Caudipteryx a Dawid Mazurek Archaeopteryx.
wtedy będzie wszystko ładnie linkowane bez przekierowania do "nie ma takiego hasła"
PS no i oczywiście Łukasz włączy ED
Ok.
EDIT: Dawid Mika postara się szybko ogarnąć Caudipteryx
Re: Archeopteryks deinonychozaurem i nomenklatura Paraves
Mnie to nie przekonuje. Sądzę, że tego typu sprawy warto ciągle testować. Mogę się mylić, ale zakładam, że wszystkie mecierze dot. celurozaurów powstałe po 2000 r. uwzględniają tą cechę. Ciekawe, co by było, gdyby pousuwać z analiz celurozaurów wszystkie cechy, które obecnie wydają się homoplastyczne. Zostaną te, które uważa się obecnie za unikalne synapomorfie. Jaki będzie kształt drzewa, trudno przewidzieć, zapewne nie będzie zbyt podobny do piertownego, na którego kształt mialy wplyw także cechy homplastyczne.Dawid Mazurek pisze:Testowanie tego, czy stopa arktometatarsalna powstała raz czy wiele razy to jedna sprawa (osobny artykuł), a filogeneza teropodów to druga. Jeżeli konsens będzie taki, że powstała raz, można jej będzie używać w pracach o filogenezie, a jak odmienny - cecha wylatuje.Jaki jest powód, dla którego uważacie, że nie powinniśmy ciągle testować - nawet wydawałoby się odrzuconych - koncepcji?
Jeżeli konsens jest taki, że powstała raz, to po co to zamieszczać ją w analizie?
Co jeśli obecny konsensus o wielokrotnym powstaniu arktometatarsala jest wynikiem przeważenia przez śmieciowe cechy, jak "jakieś przypadkowe proporcje i kształty kości kończyn i czaszki"?
Zapewne właśnie tak jest w sporej części, bo to w końcu przefenetyzowane analizy celurozaurów!Dobry przykład.Dawid Mazurek pisze:Nie kodujemy obecności piór i włosów jako tego samego stanu tej samej cechy, ale jeśli ktoś wykaże ich homologię, może będziemy musieli zacząć...
Nie kodujemy, bo mamy mocne powody (chemiczne, morfologiczne), że to nie są struktury homologiczne. Jeśli jednak okazałoby się, że to może być potencjalna synapomorfia, to myślę, że warto to wziąć pod uwagę jako cechę ewentualnie łącząca ptaki i ssaki.
Co do arktometatarsus, to nie mamy tak mocnych powodów - poza innymi cechami, co dotyczy też piór i włosów - by zakładać konwergencję (oczywiście arktometatarsus to tylko przykład, bo chodzi nam o samą zasadę).
Ale to nie będzie testowanie tej cechy.Dawid Mazurek pisze:Można zrobić nakładając tę cechę na już zrobiony bez użycia homoplazji kladogram.Powiedzmy, że na gruncie obecnych hipotez filogenezy stopa arktometatarsalna powstała 6 razy. Dlaczego jednak nie sprawdzić, czy przypadkiem 4?
Zgadza się. Ale jakie cechy są "dobre"? Te, które obecnie - na podstawie śmieciowych cech - uważamy za niehomoplastyczne.Dawid Mazurek pisze:Dokładnie. Komputerowa analiza filogenezy ma charakter statystyczny, probabilistyczny, więc używanie śmieciowych danych wejściowych może powodować sztuczne zbliżanie/oddalanie się taksonów, czego byśmy nie zaobserwowali używając "dobrych" cech.Ta cecha, zapewne przy wsparciu innych, może wszak spowodować, że pewne taksony się do siebie zbliżą, czego byśmy nie zaobserwowali, odrzucając tą cechę z góry.
Jakie to by były cechy dla celurozaurów? Pisałeś wcześniej zdaje się o kręgach, ale tutaj też jest wiele cech, które wydają się mocno narożone na homoplazje (pneumatyzacja, wysokość wyrostków itp.).
Oczywiście. Tylko tutaj wiemy, że to homplazja. Jestem jednak pewien, że bez tej wstępnej selekcji, którą proponujesz, ta cecha byłaby uważana za bardzo mocną .Dawid Mazurek pisze:Dokładnie. Cechy nie są równowartościoweWydawałoby się, że stopa arktometatarsalna to cecha, która nie powstaje w ewolucji łatwo...
Zgadza się. Wiesz, przynajmniej teoretycznie, jak działa dana metoda. Chciałbym więc poznać Twoją opinię i argumentację.Dawid Mazurek pisze:Co ktoś z nas może sądzić, a jak się w praktyce tworzy macierze i przeprowadza analizy... dwie różne sprawy.Nie sądzicie, że remedium na te problemy jest odrzucenie maximum parsimony i stosowanie innych metod, jak maximum likelihood?
Mnie natomiast przekonuje argumentacja Morimera, choć akurat to zacytowane zdanie nie, gdyż może być lepiej rozwiązana topologia (brak politomii), nie znaczy automatycznie, że wynik jest bliższy prawdy.Dawid Mazurek pisze:Podoba mi się co napisał Bennett i generalnie się pod tym podpisuję, a omawiany przez nas przypadek jaszczurek stoi w sprzeczności z tymi słowami Mortimera: "First, there have been numerous papers showing adding taxa and characters usually results in a more accurate tree". Ja bym powiedział, że dodawanie pojedynczych cech i taksonów do już istniejących macierzy (lub po prostu podobny schemat budowania nowych macierzy) z natury rzeczy zazwyczaj nie ma wystarczającej siły statystycznej by obrócić topologię o 180 stopni, więc podrasowana analiza jest zgodna w ogółach ze swoją poprzedniczką.
Z drugim zdaniem oczywiście racja.
Wcześniejszą naszą dyskusję o Scansoriopterygidae wklejam niżej (najistotniejsze kwestie). Powyższe powody pomijania Scansoriopterygidae są dla mnie niewystarczające. Z morfologią są problemy, ale dane, które oferują, należy wziąć pod uwagę, z odpowiednią ostrożnością. Rozprzestrzenienie czasowe i geograficzne oczywiście trzeba przyjmować dla znanych taksonów a kolejne mogą nie być nigdy znane. Są wczesne i z Azji, więc mogą być dobrym modelem. Owszem z macierzy wycina się niekompletne taksony, które destabilizują topologię, ale co do zasady powinno się to robić a posteriori, często a priori się usuwa fragmentaryczne (właściwie nie tyle usuwa, co nie uwzględnia a priori), ale one też mogą nieść istotne informacje.Dawid Mazurek pisze:Nie pamiętam co napisałem, ale bardziej mi chodziło o to, że póki nie za wiele wiemy o tym co to za grupa, jaka jest morfologia, pozycja systematyczna, rozprzestrzenienie czasowe i geograficzne, dla przejrzystości wywodu można je pominąć, tak jak czasem z macierzy wycina się niekompletne taksony, które destabilizują topologię... Alternatywnie: JA niewiele o niej wiem, więc ją pomijam, upraszczam, myślę syntetycznie...Dawid - wcześniej stwierdziłeś, że w swoim scenariuszu nie bierzesz pod uwagę Scansoriopterygidae, bo powinniśmy zakładać, że ich podobieństwo do owiraptorozaurów (i ogólnie 'morfotypu roślinożernego'), to konwergencja
Dawid Mazurek pisze:Podtrzymuję zdanie, że przedstawiony scenariusz z serii "a co jeśli..." był możliwy i kto wie czy nie zaszedł, choć oczywiście niekoniecznie jest to najbardziej parsymoniczny* wybór w oparciu o wspominane prace, no i z pewnością wymaga wielu założeń i uproszczeń. (...)nazuul pisze:No właśnie, stwierdzenie problamatyczności i wyspecjalizowania Scansoriopterygidae ma załatwiać sprawę. Ja bym się jednak skłaniał ku total evidence, jak widzimy poczyniłeś wiele subiektywnych/konrtowersyjnych pominięć/skrótów i ich nie wyjaśniłeś, no więc musiałem skrytykowaćDawid Mazurek pisze:Tak, to o to chodzi! Nie wziąłem ich pod uwagę! Dlaczego? Jest to grupa okryta niedawno, nieliczna, a znani przedstawiciele są stosunkowo późni czasowo w stosunku do możliwych pozycji filogenetycznych grupy. Poza tym wydają się być grupą wyspecjalizowaną!!! [convergence alert!] Informatywny filogenetycznie (bo potencjalnie wyrośnięty) jest tylko holotyp Epidexipteryx hui. Pozycja grupy u Agnolína i Novasa (2011) i Turnera, Makovicky'ego i Norella (2012) jest niestabilna i na przykład niewiele potrzeba bo zrobić z nich owiraptorozaury. Kto wie gdzie wylądują za kilka dorosłych szkieletów i za kilka macierzy... Na potrzeby mojego scenariusza, dokonałem subiektywnego wyboru, by póki co nie brać ich pod uwagęnazuul pisze:W skrócie: Twoja interpretacja jest nieprawidłowa, bo nie bierzesz pod uwagę Scansoriopterygidae.
Szczegółowo:(...)
chyba doszliśmy do meritum. Tutaj właśnie się rożnimy, gdyż moim zdaniem bardziej prawdopodobne - bo bardziej parsymoniczne - jest to, że ostatni wspólny przodek dwóch kladów był bardziej podobny do kolejnych grup zewnętrznych tego kladu, niż to, że - jak wynika z odrzuconej przez Ciebie alternatywnej hipotezy - mofologia tych grup zewnętrznych jest wynikiem konwergencji. Inaczej - wg Ciebie bardziej prawdopodobne jest to, że morfotyp archeopteryksa się pojawił u podstawy eumaniraptora)i nastąpiła rewersja u podstawy Avialae, niż to, że się pojawił u podstawy Deinonychosauria.Dawid Mazurek pisze:Moim zdaniem nieupoważnione jest założenie, że wspólny przodek dwóch kladów ma bardziej przypominać dalekich potomków wspólnego przodka z linią siostrzaną tych dwóch kladów niż po prostu bazalnych przedstawicieli któregoś z owych dwóch kladów.
zobrazować to można w ten sposób, że "morfotyp archeopteryksa" to A natomias B to "morfotyp rośniożerny" (grup zewnęrznych do Deinonychosauria wg Godefriota i in., cechy za Xu i in. 2011- oczywiście wiemy, że to jest katasfrofalne uproszczenie i dobór cech i inne parametry są tu kluczowe). Mamy więc coś takiego, gwiazdką oznaczyłem formy jurajskie:
B(B*(A*(B+B)))
są to po kolei:
Incisivosaurus/Protaracheopteryx (=bazalne Oviraptorosauria), Scansorioprerygidae, (tu mamy deinonychozaury - A), Shenzhouraptor, Omnivoropterygidae
Wg Ciebie "morfotyp A" to pierwotna budowa podkreślonego kladu:
B(B*(A*(B+B)))
a wg mnie tego:
B(B*(A*(B+B)))
Błąd widzę u Ciebie w tym, że nie bierzesz pod uwagę Scansoriopterygidae - B* (EDIT - jeszcze coś - patrz pozycje Epidexipteryx i Epidendrosaurus w Turner i in. 2012).
Skutkuje to tym, że "morfotyp A" uważasz za pierwotny dla Avialae:Oczywiście u podstawy Eumaniraptora musiały nastąpić zmiany w układzie lokomocji (kończyny przednie) ale zakłądam, ze morfotyp archeopteryksa to coś więcej, m.in. cechy czaszkiDawid Mazurek pisze:Moim zdaniem z kladogramu Godefriota i in. wynika jasno, że bazalny deinonychozaur musiał być Archie-like, a to że taki morfotyp mógł mieć też bazalny Avialae domyśliłem już sam, ale w zgodzie z licznymi scenariuszami wywodzącymi przez dziesięciolecia ptaki od archeopteryksa.
* Swoją drogą, problem parsymonii jest w przypadku macierzy morfologicznych bardzo problematyczny. Wiemy, że dla ustalania pokrewieństw liczą się tylko synapomorfie, nigdy autapomorfie. O ile w analizach molekularnych używa się śmieciowych sekwencji, na które nie działa presja selekcyjna, to w morfologii robi się coś przeciwnego (nie moja obserwacja, wyszło wczoraj w pewnej dyskusji)! Jeżeli dwa taksony mają wspólne cechy związane z budową kończyn, zębów, proporcjami elementów czaszki, to biorąc pod uwagę POWSZECHNOŚĆ konwergencji i znaną wartość adaptatywną tych elementów szkieletowych, oraz ograniczoną liczbę możliwych morfotypów w morfoprzestrzeni przystosowań funkcjonalnych i ekologicznych, czy nie bardziej parsymoniczne jest brać jako hipotezę 0, że taka spójność wskazuje na konwergencję? Może program powinien takie taksony, szczególnie jeżeli pochodzą z różnych horyzontów czasowych, domyślnie umieszczać na kladogramie osobno jako coś oszczędniejszego? ;-) My zaś przy odtwarzaniu filogenezy, zamiast zajmować się całym szkieletem, powinniśmy się skupiać na... bo ja wiem... kręgach, histologii... kontekście czasowym i biogeograficznym. Udało się to wyłapać w przypadku odległych konwergencji, np. popozaury i ornitomimozaury, ale w obrębie samych dinozaurów, wiele stanów, adaptacji mogło powstawać niezależnie, częste rewersje, paralelizmy, ewolucja iteratywna... myślę że BARDZO tego nie doceniamy i zastanawiam się na ile nasze klarowne obrazy filogenezy są jednym wielkim bajzlem, a wiele 'synapomorfii' to homoplazje
Trafne stwierdzenia, z pewnością warte bliższego rozważenia.Dawid Mazurek pisze:No skoro tak, to faktycznie dużo to zmienia, a więc pozycja w obrębie owiraptorozaurów, lub informatywna dla badania początków większego kladu (np. Paraves+Oviraptorosauria) plezjomorficzna budowa, być może ujawniona w związku z wiekiem ontogenetycznym, kto wie, może wręcz 2/3 Scansoriopterygidae to po prostu polifiletyczna grupa pisklaków? To by pasowało!!! Wiemy też, że wszystkie 3 okazy mogą być synonimiczne - dwa pisklaki i jeden dorosły o cechach pisklaków/przodków. Tak tylko głośno myślę... Innych możliwości nie widzę, no chyba że rekonstrukcje czaszek są tendencyjne...Jak już wcześniej wskazałem w tym wątku, to może być plezjomorficzna budowa Paraves+Oviraptorosauria (Xu i in. 2011, Xiaotingia).
Bardzo podobny wydaje się też Sapeornis (którego Paul włączył do Oviraptorosauria!)
Alwarezzauroidy czy terinozozaury spokojnie można pominąć, gdyż jest spora szansa, że są bardziej bazalne.Dawid Mazurek pisze:Nie chce mi się bazgrać, więc tak w skrajnym uproszczeniu: późna jura - najstarsze znane ptaki (linia Archaeopteryx i Anchiornis) współwystępują z pierwszymi wtórnymi nielotami (Scansoriopterygidae i ich owiraptorozaurzy potomkowie). Wczesna kreda - druga fala wtórnych nielotów (tetrapteryksy) i dalej z nich troodony, z których z kolei w później kredzie dromeozaury. Gdyby taki scenariusz był prawdziwy (a wątpię, aż tak prosto nie ma), to stosując definicje musielibyśmy powiedzieć, że troodony i dromeozaury powstały gdzieś w środkowej kredzie, a wcześniejsze 'troodony' to w rzeczywistości bazalne deinonychozaury. Ja wolę jednak by taksonomia nie pomijała aspektów anatomicznych i powiedziałbym że przy takim scenariuszu dromeozaury wydzieliły się z troodonów, które trwały nadal. Terizinozaury i alwarezzaurydy? Nie wiem, nie mam pomysłu na tą chwilę, musiałbym polookać więcej w drzewka...Też mam czasem wrażenie, że kladogramy są na odwrót.
Mógłbyś przedstawić proponowaną przez siebie topologię?
Nie musisz bazgrać, wystarczy, że napisałbyś coś takiego (jeśli dobrze odczytuję):
Archaeopteryx, Anchiornis [itp.] (Scansoriopterygidae+Oviraptoridae) (Microraptoria [?Sinovenator, Mei] (Troodontidae+Dromaeosauridae))
nie wiadomo, gdzie mają być problmatyczne Unenlagiinae i Rahonavis. Powyższy scenariusz powoduje, że z trzech głównych ghost lineages (do Troodontidae, Dromaeosauridae i Microraptoria) powstaje jedna, czyli tak jak w większości hipotez, gdzie Microraptoria należy do Dromaeosauridae, które są taksonem siostrzanym Troodontidae.
Moim zdaniem znacznie przeceniasz znaczenie rozmieszczenia czasowego i przestrzennego znanych form. Oczywiście zawsze można stwierdzić, że zapis kopalny jest niekompletny, ale w tym przypadku jest to dobrze uzasadnione. Tak złożyło, że znamy kilka taksonów z wąskiego przedziału czasowego ze środkowej-późnej jury Chin (?oksford), jeden z końca jury Niemiec (tyton), później długo nic (oczywiście są ząbki, ale one są bardzo podejrzane) i od wczesnej kredy (?barrem) bardzo wiele, zwłaszcza z Chin i USA.
Anchiornis, Xiaotingia i Eosinopteryx znane są praktycznie z tych samych osadów, zbliżone są Pedopenna, Epidexipteryx i pisklaki Epidendrosaurus (=Scansoriopteryx). Pierwsze trzy oczywiście mogą być bazalnymi przedstawicielami kladu, który dał początek troodontydom, dromeozaurydom, ptakom, ale jak najbardziej też mogą być przedstawicielami jednej z tych linii (choć one same niekoniecznie tworzą odrębny klad) - właśnie z uwagi na to, że są tak bliskie czasowo i geograficznie. Biorąc np. topologię Agnolina i Novasa, to, że z tych samych osadów nie są znane linie bardziej bazalne niż te, które tam są obecne, nie znaczy, że nie było ich gdzieś indziej. Z Tiaojishan podobno znany jest alwarezzauroid podobny do Haplocheirus (takie plotki gdzieś w necie krążyły, nie wiem na ile to prawda). Nie ma natomiast tyranozauroidów czy ceratozaurów, które znane są z pobliskich osadów (Guanlong, Limusaurus), nie ma też przedstawicieli wielu innych grup, które powinny być tam obecne, jest za to sporo pterozaurów i tianyulongów. Jak dla mnie, to zdecydowanie za słabo znamy Tiaojishan i okolice, żeby z tego wyciągać daleko idące wnioski o niezgodności hipotezy filogenetycznej z czasem i geografią. Z Tiaojishan, Jehol czy Solnhofen znamy dużo małych taksonów, natomiast z innych stanowisk jest ich mało. Z Jehol znane są podobno setki(!) mikroraptorów, później długo nic, ale występowały też pojedyncze sztuki w kampanie-mastrychcie (Hesperonychus, Luanchuanraptor), które znane są z dobrze przeczesanych osadów. Czy to znaczy, że to nie są mikroraptory? Inna sprawa to formy gondwańskie, które mają najdłuższe ghost lineages. Oczywiście Rahonavis jako relikt, bardziej bazalny od zapewne wszystkich ptaków z Jehol czy Hiszpanii jest mało przekonujący, ale na ile znamy skały, gdzie szczątki jego przodka mogłyby być obecne? Na ile znamy w ogóle jurajskie i kredowe Paraves z Afryki, Madagaskaru, Indii? Tak samo Unenlagiinae. Sądzę, że filogenezę form gondwańskich trzeba uznać za bardzo niepewną i tu się pewnie zgodzisz (na podstawie anatomii, o której piszą Agnolin i Novas, wydaje się prawdopdobne, że to formy wtórnie naziemne).
Warto też wziąć pod uwagę 'Lori' - nienazwanego troodontyda z późnej jury Ameryki Płn., opisanego tylko w posterze z konferencji SVP z 2005 r. ( http://www.bhbfonline.org/AboutUs/Lori.pdf ). Polecam przeczytać, pewne dane są bardzo interesujące.
Bardzo zaawansowane dromeozaurydy są już obecne we wczesnej kredzie - ?barrem (Utahraptor).Dawid Mazurek pisze:Wczesna kreda - druga fala wtórnych nielotów (tetrapteryksy) i dalej z nich troodony, z których z kolei w później kredzie dromeozaury. (...) stosując definicje musielibyśmy powiedzieć, że troodony i dromeozaury powstały gdzieś w środkowej kredzie
Odnalezione w Nigrze tropy zostały pozostawione przez duże (3-4 m, wg Xinga i in., [2012] aż 4,7 m), prawdopodobnie środkowojurajskie deinonychozaury (Mudroch i in., 2011).
Mudroch, A., Richter, U., Joger, U., Kosma, R., Idé, O. & Maga, A. (2011) "Didactyl Tracks of Paravian Theropods (Maniraptora) from the ?Middle Jurassic of Africa" PLoS ONE 6(2), e14642. doi:10.1371/journal.pone.0014642
Xing, L., Li, D., Harris, J.D., Bell, P.R., Azuma, Y., Fujita, M., Lee, Y.-N. & Currie, P.J. (2012) "A new Dromaeosauripus (Dinosauria: Theropoda) ichnospecies from the Lower Cretaceous Hekou Group, Gansu Province, China" Acta Palaeontologica Polonica, doi:10.4202/app.2011.0115
Które cechy obecne u Troodontidae, są tak istotne, żeby dla samej tej morfologii robić z nich grad i które cechy dromeozaurydów są tak istotne, że odróżniają je od troodontydów?Dawid Mazurek pisze:Ja wolę jednak by taksonomia nie pomijała aspektów anatomicznych i powiedziałbym że przy takim scenariuszu dromeozaury wydzieliły się z troodonów, które trwały nadal
Inaczej mówiąc - czym jest morfologia troodontydowa i dlaczego jest aż tak ważna dla nomenklatury?
Zmieniać scenariusz, ale nie omijać wcześniejszych danych.Dawid Mazurek pisze:Nie tylko o to mi chodzi. Zarzuciłeś któremuś z moich pomysłów, że nie bierze pod uwagę Scansoriopterygidae. Ja w takim momencie mówię: poczekajmy, będzie 10 gatunków i konsens co do ich pozycji filogenetycznej, weźmie się je pod uwagę. Nie ma co się trzymać ustaleń sprzed roku, bo co kladogram trzeba by zmieniać scenariusz.Rozumiem, że chodzi o to, że jestem zwolennikiem nomenklatury filogenetyczej i metody badania filogenezy za pomocą numerycznych analiz filogenetyczych. Nie wydaje mi się, żeby to była kwestia przyjęcia przeze mnie aktualnej mody... Mógłbyś podać konkretne przykłady, które potwierdzają Twoje słowa?
Zarzuciłem to scenariuszowi przedstwionemu przez Ciebie na samym początku wątku, który wyraźnie opiera się na topologii Godefroit et al. (2013).
Co jeśli nie będzie nowych gatunków ani konsensusu do pozycji? Te pierwsze jest zupełnie nieistotne a drugiego może też nigdy nie być. Te argumenty mnie więc totalnie nie przekonują. Jak już stwierdziłem, wolę total evidence.
Na dzień dzisiejszy można uznać Scansoriopterygidae za Maniraptora incertae sedis - mogą być u podstawy: Avialae, Ornithurae, Paraves, Oviraptorosauria, obok Shenzhouraptor a nawet w Alvarezsauroida. Kto wie, może w niektórych hipotezach będą w Therizinosauria, Omnirovopterygidae albo Deinonychosauria - z tymi dwoma pierwszymi mają pewne wspólne cechy a co do Deinonychosauria, to mogą nawet reprezentować ich pierwotną morfologię (zob. np. Xu i in. 2011). Możliwe, że Scansoriopterygidae reprezentują pierwotną morfologię Maniraptora. Taki pogląd na systematykę - Maniraptora i.s. - będzie zapewne aktualny jeszcze długo. Ponieważ z dużą dożą prawdopodobieństwa znajdują się blisko podstawy Paraves i są stare (do czego przywiązujesz dużą wagę), to, choć są problematycze, należy je wziąć szczególnie pod uwagę, nawet samego Epidexipteryx, może nawet tym bardziej, że mogą być plezjomorficzne także przez młodocianość.
Nie mam zielonego pojęcia, ile może trwać powstanie akywnego lotu u np. lotopałanek. Może kilkanaśie milionów lat? Były jakieś badania na temat, ile może/mogło trwać powstanie lotu?Dawid Mazurek pisze:Daję ewolucji czas na zrobienie archeopteryksa, biorę pod uwagę prawdopodobną ghost lineage (szybujący przodek archeopteryksa z jakichś lasów na archipelagu wysp z którego nic w zapisie kopalnym nie przetrwało; pomyśl o rozprzestrzenieniu geograficznym dzisiejszych organizmów z perspektywami na aktywnie latających potomków), no i problematyczne jeżeli chodzi o datowanie ptasie (lub nie ptasie) tropy.Dlaczego wczesna jura czy trias?
Mógłbyś dać jakiaś źródło twierdzenia, że na tym archipelagu były lasy?
O ile się nie mylę, nie ma tak wczesnych tropów, które pochodzą na pewno od ptaka (nie za bardzo rozumiem, co jaki klad chodzi, ale chyba o zawierający archeopteryksa; Martyniuk 2012 utworzył fajną nazwę Ornithes dla kladu archeopteryks+crown Aves).
z archozaurami sprawa jest bardziej skomplikowana, choć obecnie raczej ogranicza się je do ptaków+krokodyliDawid Mazurek pisze:No gdyby tak było, że specyfikatorami w przypadku archozaurów byłyby i jakiś fitozaur i jakiś aetozaur i jakiś dinozaur i... to OKokreślony zestaw taksonów = specyfikatory
Kwestia podejścia - do mnie bardziej przemawia kilka dobrze zdefiniowanych nazw i drzewko, bez zastanawiania się, czy dana nazwa dotyczy grupy monofiletycznej czy gradu, choć się zgadzamy, że co za dużo to niezdrowo.Dawid Mazurek pisze:Jak topologie drzew się ustabilizują, to znajdą się ludzie, którzy zaczną nazywać każde odgałęzienie, powstanie kupa nowych nazw, których nikt nie spamięta i po samej nazwie nie będzie w stanie powiedzieć, czy to jakaś ważna grupa, stąd jestem za tworzeniem jednostek parafiletycznych i nadawanie im rang. W przypadku trzech kolejnych taksonów leżących coraz bardziej w stronę korony (A(B(C(D-Z)))), lepiej utworzyć dla nich jedną jednostkę o randze mówiącej że to SUBIEKTYWNIE mała grupa, czyli zrobić rodzinę dla A, B i C, niż nazywać każdy z kladów z osobna.Myślę że w ostatnich kilku latach w nieptasiodinozaurologii jest tylko kilka sensownych propozycji nowych nazw, reszta jest niepotrzebna.
-
d_m
Re: Archeopteryks deinonychozaurem i nomenklatura Paraves
Póki co napiszę tylko, że generalnie zgadzam się w materii faktów, rozbieżności są paradoksalnie niewielkie i ograniczone do sfery podejścia filozoficznego do problematyki taksonomii i nomenklatury. Wynika to z tego, że przypisujemy inną rangę zastrzeżeniom i założeniom leżących po obu stronach podejścia, o których to istnieniu obaj dobrze wiemy. Dodam, że do żadnego konkretnego scenariusza ewolucyjnego powstania ptaków nie jestem jakoś specjalnie przywiązany. W kilku kwestiach zasób wiedzy nie pozwala mi za wiele odpisać, ale jak się trochę dokształcę to spróbuję. Ten las na archipelagu to hipoteza rzucona ad hoc na potrzeby dyskusji, odnosząca się do przodków ptaków i nie miała się kojarzyć z siedliskiem życia Archaeopteryx. Ciekawa ta informacja o Lori.
Znajomy do mnie pisał z pytaniem o jaką prackę chodzi w newsie. Może poza linkiem do forumowej dyskusji, warto by było do newsa wstawić też cytację i DOI-link książki.
Znajomy do mnie pisał z pytaniem o jaką prackę chodzi w newsie. Może poza linkiem do forumowej dyskusji, warto by było do newsa wstawić też cytację i DOI-link książki.
-
Ag.Ent
- Kredowy tyranozaur
- Posty: 2349
- Rejestracja: 19 marca 2009, o 20:55
- Imię i nazwisko: Tomasz Skawiński
- Lokalizacja: Wrocław
Re: Archeopteryks deinonychozaurem i nomenklatura Paraves
Dobry pomysł. Żeby już się nikt nie męczył (poza tym mam dziwne upodobanie do tworzenia cytacji):Dawid Mazurek pisze:Znajomy do mnie pisał z pytaniem o jaką prackę chodzi w newsie. Może poza linkiem do forumowej dyskusji, warto by było do newsa wstawić też cytację i DOI-link książki.
Można jeszcze wstawić zdjęcie okładki, chociaż jest brzydkaAgnolín F.L., Novas F.E. 2013. Avian Ancestors. A Review of the Phylogenetic Relationships of the Theropods Unenlagiidae, Microraptoria, Anchiornis and Scansoriopterygidae. Springer, ss. 96. ISBN 978-94-007-5637-3, http://dx.doi.org/10.1007/978-94-007-5637-3.
Tak wiele dinozaurów, tak mało czasu...
-
d_m
Re: Archeopteryks deinonychozaurem i nomenklatura Paraves
To samo pomyślałem jak ją zobaczyłem. Kogo oni tam zatrudniająAg.Ent pisze:Można jeszcze wstawić zdjęcie okładki, chociaż jest brzydka
Re: Archeopteryks deinonychozaurem i nomenklatura Paraves
Istotna dla dyskusji praca o tylnych skrzydłach bazalnych ptaków:
Zheng et al. 2013 Hind Wings in Basal Birds and the Evolution of Leg Feathers, Science, http://dx.doi.org/10.1126/science.1228753

Sapeornis autorstwa Yike Xu, źródło: http://www.history.com/news/did-four-wi ... ient-skies
EDIT
Zheng et al. 2013 Hind Wings in Basal Birds and the Evolution of Leg Feathers, Science, http://dx.doi.org/10.1126/science.1228753
Sapeornis autorstwa Yike Xu, źródło: http://www.history.com/news/did-four-wi ... ient-skies
EDIT
Zheng et al. 2013, suplement pisze:Functions of the leg feathers in basal paravians. It is widely accepted that the large leg feathers of Microraptor were used in aerial locomotion, but no consensus has been reached as to how these feathers conferred an aerodynamic advantage (1, 5, 6, 10, 24, 25, 49, 50). Few studies have even addressed this question with regard to other basal paravians. Such morphological features as asymmetrical vanes and curved, robust rachises may be associated with flight, but in no case has this kind of association been strictly demonstrated. For example, the presence of vane asymmetry and curvature has been suggested to have an aerodynamic function (51), but both features are also present in the feathers of at least some flightless birds (52, 53). However, it is still possible to infer the functions of particular feather types based on a synthetic overview of the available data.
The leg feathers of basal paravians probably differed in primary function from the metatarsal feathers that are seen in some modern birds. The latter are fluffy and oriented nearly parallel to the long axis of the metatarsus, and have been suggested to function mainly in insulation or protection (31). The leg feathers of basal paravians are considerably different from modern examples in both general morphology and arrangement, and appear less likely to have served primarily as insulation. It is also unlikely that basal paravian leg feathers were primarily for display. In living birds, any large feathers that have a display function and are not also used in flight are normally located on the head or tail, and they differ from flight feathers in lacking such features as vane asymmetry and shaft curvature. The leg feathers of basal birds and other basal paravians have curved rachises, are nearly perpendicular to the long axis of the leg, and are arranged in a wing-like manner (forming a planar surface). These features suggest that
basal paravian leg feathers are more likely to have been aerodynamic in primary function, perhaps creating lift and/or enhancing maneuverability (1, 5, 6, 10, 24, 25, 49, 50), than to have been used primarily for display or insulation. However, it is possible that the leg feathers of basal paravians combined a primary role in flight with a secondary function such as display.
Ed Young, [url]http://phenomena.nationalgeographic.com/2013/03/14/the-rise-and-fall-of-four-winged-birds/[/url] pisze:Xu thinks that these feathers might have helped the owners to fly. They could have produced extra lift or maybe helped the birds to turn more easily. But other scientists who work on the evolution of flight are not convinced. “[Xu] has basically just taken a punt that because the feathers were stiff, they were probably aerodynamic in function,” says Michael Habib from the Johns Hopkins Medical Institute. “It is a bit of a weak argument.”
Habib thinks that the long asymmetric leg feathers of Microraptor probably did play some role in gliding or flying, but the smaller plumes of other baggy-legged species “might have merely been there because of a developmental quirk”. If some genes are producing large feathers on the front limbs, “it might not take much to tweak a set onto the hind limbs too,” he says.
Kevin Padian from the University of California, Berkeley agrees. He points out that no one has actually done any proper tests to show if the leg feathers were involved in flight. They would certainly have created drag, but they could only have provided lift if they sat in a flat sheet like the wings of modern birds. Xu claims that they were, but Padian says that the feathers could just have been flattened into a plane as they became fossilised. “It hasn’t been shown that this is really an aerodynamically competent wing,” he says.
(...)
Why did the leg feathers, having first become large, eventually disappear? Xu thinks that it was because the birds set their two pairs of limbs towards different ends—the front pair for flying and the hind pair for walking or running. At the same time, they might have moved from life in the trees to life on the ground, or near water. Under all these scenarios, long leg feathers would have just got in the way, and were soon lost.
Something similar may have happened in other flying animals. For example, the earliest flying insects tend to have four wings, while some of the most competent flyers like, well, flies, only have two. The second pair has evolved into a pair of gyroscopes called halteres. “In the early evolution of flight, different animal groups always try to use as much surface as possible,” says Xu. “Once the major flight organ is well developed, the animal just fires the other organs.”
-
d_m
Re: Archeopteryks deinonychozaurem i nomenklatura Paraves
Ta publikacja pokazuje, że się zauważa tego, czego się nie szuka - pamiętam, że jednemu z archeopteryksów wręcz odpreparowano tylne skrzydła. Zmienił się paradygmat, to i odpowiednie okazy się znalazły...
Online ukazała się też pracka Preservation of ovarian follicles reveals early evolution of avian reproductive behaviour.Nie czytałem, ale z doniesień i komentarzy wynika, że identyfikacja widocznych struktur jest bardzo pobieżna. Ale co tam... jak coś ma pióra i jest z Jehol to zaraz Nature  No ale w sumie nie mam co narzekać, bezkręgowce kambru też są medialne...
No ale w sumie nie mam co narzekać, bezkręgowce kambru też są medialne...
Przeczytałem The evolution of the feather: scales on the tail of Sinosauropteryx and an interpretation of the dinosaur’s opisthotonic posture pióra Lingham-Soliara (2013) - napisane ciekawie i przekonująco, kilka kluczowych uwag i ważnych informacji się tam znajduje, ale... materiał dowody beznadziejnie zilustrowany, więc wnioski niewiarygodne! Czekam na pracę w przygotowaniu o rozkładzie młodych strusi...
Online ukazała się też pracka Preservation of ovarian follicles reveals early evolution of avian reproductive behaviour.
Przeczytałem The evolution of the feather: scales on the tail of Sinosauropteryx and an interpretation of the dinosaur’s opisthotonic posture pióra Lingham-Soliara (2013) - napisane ciekawie i przekonująco, kilka kluczowych uwag i ważnych informacji się tam znajduje, ale... materiał dowody beznadziejnie zilustrowany, więc wnioski niewiarygodne! Czekam na pracę w przygotowaniu o rozkładzie młodych strusi...
-
d_m
Re: Archeopteryks deinonychozaurem i nomenklatura Paraves
Hmmm... jesteśmy w stanie dodać Haplocheirus do macierzy Agnolína i Novasa (2013) i zobaczyć co się stanie? To niby ponad 400 cech, ale z Science aż tyle się ich nie wyciągnie, a rąk do roboty może by się kilka znalazło...
-
Ag.Ent
- Kredowy tyranozaur
- Posty: 2349
- Rejestracja: 19 marca 2009, o 20:55
- Imię i nazwisko: Tomasz Skawiński
- Lokalizacja: Wrocław
Re: Archeopteryks deinonychozaurem i nomenklatura Paraves
Być w stanie to może i jesteśmy, ale to jednak strasznie dużo mało przyjemnej roboty. Może pamiętacie, że próbowałem swego czasu zakodowaćDawid Mazurek pisze:Hmmm... jesteśmy w stanie dodać Haplocheirus do macierzy Agnolína i Novasa (2013) i zobaczyć co się stanie? To niby ponad 400 cech, ale z Science aż tyle się ich nie wyciągnie, a rąk do roboty może by się kilka znalazło...
Re: Archeopteryks deinonychozaurem i nomenklatura Paraves
Jaki by miał być cel tego?
Pewnie, że można tak zrobić. Samego haplocheira raczej nie ma sensu, bardzo istotne będą też Tianyuraptor, Mahakala, Jinfengopteryx, Eosinopteryx i Nqwebasaurus, a najlepiej też inne (Luanchuanraptor, Pedopenna, Yixianosaurus, Balaur, Similicaudipteryx, Aniksosaurus, Ningyuansaurus, Pyroraptor, Xixiasaurus).
Tylko inni zrobią to lepiej, np. Mortimer w analizie, która ma wyjść wraz z 'Lori'.
Co do wcześniejszej dyskusji:
Nie wiem na ile trafne jest spostrzeżenie, że ludzie zajmujący się morskimi bezkręgowcami, które są stosunkowo prosto zbudowane a zapis kopalny mają często świetny, przekładają założenia i metody dobrze działające na tym gruncie na lądowe kręgowce, które są bardzo skomplikowane a zapis jest często tragiczny (np. dla dyskutowanych tu bazalnych Maniraptora).
Pewnie, że można tak zrobić. Samego haplocheira raczej nie ma sensu, bardzo istotne będą też Tianyuraptor, Mahakala, Jinfengopteryx, Eosinopteryx i Nqwebasaurus, a najlepiej też inne (Luanchuanraptor, Pedopenna, Yixianosaurus, Balaur, Similicaudipteryx, Aniksosaurus, Ningyuansaurus, Pyroraptor, Xixiasaurus).
Tylko inni zrobią to lepiej, np. Mortimer w analizie, która ma wyjść wraz z 'Lori'.
To zmora tego typu publikacjiDawid Mazurek pisze:Przeczytałem The evolution of the feather: scales on the tail of Sinosauropteryx (...) materiał dowody beznadziejnie zilustrowany, więc wnioski niewiarygodne!
Co do wcześniejszej dyskusji:
Też mi się tak wydaje.Dawid Mazurek pisze:Póki co napiszę tylko, że generalnie zgadzam się w materii faktów, rozbieżności są paradoksalnie niewielkie i ograniczone do sfery podejścia filozoficznego do problematyki taksonomii i nomenklatury. Wynika to z tego, że przypisujemy inną rangę zastrzeżeniom i założeniom leżących po obu stronach podejścia, o których to istnieniu obaj dobrze wiemy.
Nie wiem na ile trafne jest spostrzeżenie, że ludzie zajmujący się morskimi bezkręgowcami, które są stosunkowo prosto zbudowane a zapis kopalny mają często świetny, przekładają założenia i metody dobrze działające na tym gruncie na lądowe kręgowce, które są bardzo skomplikowane a zapis jest często tragiczny (np. dla dyskutowanych tu bazalnych Maniraptora).
-
d_m
Re: Archeopteryks deinonychozaurem i nomenklatura Paraves
Chociaż opisywano w świecie inne stare "ptasie" tropy (min. Plesiornis z Polski), można uznać, że sztandarowy przykład, którym się zasugerowałem, został obalony przez głównego autora pierwotnego doniesienia.Dawid Mazurek pisze:Daję ewolucji czas na zrobienie archeopteryksa, biorę pod uwagę prawdopodobną ghost lineage (szybujący przodek archeopteryksa z jakichś lasów na archipelagu wysp z którego nic w zapisie kopalnym nie przetrwało; pomyśl o rozprzestrzenieniu geograficznym dzisiejszych organizmów z perspektywami na aktywnie latających potomków), no i problematyczne jeżeli chodzi o datowanie ptasie (lub nie ptasie) tropy.nazuul pisze:Dlaczego wczesna jura czy trias?
1) Bird-like fossil footprints from the Late Triassic. Nature 417, 936-938 (27 June 2002)
http://www.nature.com/nature/journal/v4 ... 00818.html
2) A Late Eocene date for Late Triassic bird tracks. Nature 495, E1–E2 (21 March 2013)
http://www.nature.com/nature/journal/v4 ... 11931.html
-
d_m
Re: Archeopteryks deinonychozaurem i nomenklatura Paraves
Niedawno zmarł BAND(Birds Are Not Dinosaurs)yta Larry Martin, który w późniejszych latach uważał (niektóre?) maniraptory za wtórnie nielotne ptaki, pogląd z którym na dzień dzisiejszy się zgadzam. Kilka akapitów o jego działalności:
http://jasonbrougham.com/2013/03/31/mem ... ry-martin/
http://jasonbrougham.com/2013/03/31/mem ... ry-martin/
Re: Archeopteryks deinonychozaurem i nomenklatura Paraves
Często zapomina się o tym, że BAND to równoprawna wobec BAD hipoteza. Wprawdzie BAD opiera się na wielu danych, ale jako hipoteza wiodąca nie jest wystarczająco testowana, np. praktycznie wszystkie analizy filogenetyczne z góry zakładają ten scenariusz, bo brak w nich np. longiskwamy. Np. omawiane tu prace Dececchi & Larsson, 2011 czy Agnolin & Novas, 2013 nie mają wielkiej wartości z tego punktu widzenia.
Możliwe, że to, co uważa się powszechnie za bardziej zaawansowane od archeopteryksa ptaki to prawdziwe ptaki, które pochodzą od longiskwamy, a to, co uważa się za linię dinozaurów wiodącą ku ptakom, to dinozaury konwergentnie zbliżające się do budową do ptaków (np. archeopteryks jest bardzo podobny do deinonychozaurów). Możliwe, że ptaki są difiletyczne.
EDIT 2.4.2013
Post napisałem 1. kwietnia, co wpływa na jego treść. Pierwszy akapit wskazuje na rzeczywiste zjawisko. Drugi akapit przedstawia wprawdzie prawdę ("możliwe, że"), ale taki scenariusz jest bardzo nieoszczędny, biorąc pod uwagę wszystkie dane i dlatego należy go odrzucić.
Możliwe, że to, co uważa się powszechnie za bardziej zaawansowane od archeopteryksa ptaki to prawdziwe ptaki, które pochodzą od longiskwamy, a to, co uważa się za linię dinozaurów wiodącą ku ptakom, to dinozaury konwergentnie zbliżające się do budową do ptaków (np. archeopteryks jest bardzo podobny do deinonychozaurów). Możliwe, że ptaki są difiletyczne.
EDIT 2.4.2013
Post napisałem 1. kwietnia, co wpływa na jego treść. Pierwszy akapit wskazuje na rzeczywiste zjawisko. Drugi akapit przedstawia wprawdzie prawdę ("możliwe, że"), ale taki scenariusz jest bardzo nieoszczędny, biorąc pod uwagę wszystkie dane i dlatego należy go odrzucić.
-
Ag.Ent
- Kredowy tyranozaur
- Posty: 2349
- Rejestracja: 19 marca 2009, o 20:55
- Imię i nazwisko: Tomasz Skawiński
- Lokalizacja: Wrocław
Re: Archeopteryks deinonychozaurem i nomenklatura Paraves
Dobrze, że to dodałeś, bo już szykowałem się do przydługiej kontry ;), ale mimo wszystko jednak wypowiem się w kilku kwestiach.nazuul pisze:EDIT 2.4.2013
Post napisałem 1. kwietnia, co wpływa na jego treść. Pierwszy akapit wskazuje na rzeczywiste zjawisko. Drugi akapit przedstawia wprawdzie prawdę ("możliwe, że"), ale taki scenariusz jest bardzo nieoszczędny, biorąc pod uwagę wszystkie dane i dlatego należy go odrzucić.
Rozpatrując sprawę możliwie najbardziej ogólnie, to faktycznie BAD i BAND są równoprawnymi hipotezami (tzn. przynajmniej teoretycznie każda z nich może okazać się prawdziwą). Ale mimo wszystko ja bym ich takimi nie nazywał - dane, które obecnie posiadamy, wspierają tylko jedną z nich.
I wydaje się, że obecnie nie ma nawet za bardzo możliwości przedstawienia naukowej alternatywy dla BAD. Longisquama? Scleromochlus? Sfenozuchy? Ojej... Między tymi domniemanymi ptasimi przodkami a pierwszymi znanymi ptakami jest co najmniej 50 mln lat przerwy, a nie znamy żadnych, choćby potencjalnych, form pośrednich.
Tak wiele dinozaurów, tak mało czasu...