


Jesteś w: Artykuły o teropodach
Ceratozaury (Ceratosauria) bazalne
Autor: Maciej Ziegler, korekta: Tomasz Sokołowski
Ostatnia aktualizacja: 12.07.2011 r.
Nomenklatura:
- Ceratosauria (Marsh, 1884)
>Ceratosaurus nasicornis ~ Passer domesticus (zmodyfikowano z Rowe, 1989)
Do tak definiowanego kladu mogą należeć - tutaj omówione osobno - celofyzoidy. Jeżeli ta hipoteza okaże się bardziej prawdopodobna, to niniejszy artykuł dotyczy kladu Ceratosauroidea (>Carnotaurus sastrei ~ Coelophysis bauri; zmodyfikowano z Sereno, 1998). Na potrzeby tego i innych artykułów przyjmujemy dominującą obecnie - wspartą m. in. nowymi danymi nt. Majungasaurus i Limusaurus hipotezę o węższym zasięgu Ceratosauria (ale zob. też Tykoski, 2005 i Mortimer, online), którego taksonem siostrzanym jest Tetanurae; z kolei Coelophysoidea jest taksonem siostrzanym Ceratosauroidea - do którego należą (obok np. Zupaysaurus wg Smitha i in., 2007) Dilophosauridae i Averostra sensu Ezcurra i Cuny, 2007 (<Ceratosaurus nasicornis & Allosaurus fragilis - czyli tetanury i ceratozaury). W takim wypadku zasięg kladu Ceratosauroidea obejmuje niemal wszystkie teropody, dlatego nazwa ta nie jest używana dalej. - Neoceratosauria (Novas, 1992)
<Ceratosaurus nasicornis & Abelisaurus comahuensis (zmodyfikowano z Holtz, 1994) - Ceratosauridae (Marsh, 1884)
>Ceratosaurus nasicornis ~ Abelisaurus comahuensis (zmodyfikowano z Rauhut, 2004) - Abelisauroidea (Bonaparte i Novas, 1985 sensu Bonaparte, 1991)
<Abelisaurus comahuensis ~ Ceratosaurus nasicornis (Tykoski i Rowe, 2004; zmodyfikowano z Holtz, 1994) - Abelisauria (Novas, 1992)
<Abelisaurus comahuensis & Noasaurus leali (Novas, 1997)
Charakterystyka:
Ceratozaury ("rogate jaszczury", od typowego rodzaju Ceratosaurus) to takson siostrzany tetanurów, pozostający przez niemal cały mezozoik w ich cieniu. Wydaje się, że dominowały w późnej kredzie w Gondwanie i Europie, w których jednak konkurencją były (przynajmniej w Ameryce Południowej, Australii i Europie) tetanury - szczególnie karnozaury, ale także celurozaury i spinozaurydy.
Wiele bazalnych ceratozaurów (Deltadromeus, Spinostropheus, Elaphrosaurus, Limusaurus) było dość lekko zbudowanych - miały długie nogi i szyje, co upodabniało je do celofyzoidów. Jeden z nich (Deltadromeus) jest największym tak "delikatnie" zbudowanym teropodem. Inne, jak Ceratosaurus i abelizaurydy były masywne i polowały na dużą zdobycz. Niemal wszystkie ceratozaury były mięsożerne i uzbrojone w ostre zęby. Wyjątkiem jest Limusaurus - niewielki, wyspecjalizowany roślinożerca z późnej jury Chin. Wiele ceratozaurów wykazuje duże podobieństwo do przedstawicieli innych grup - generalnie są one podobne do celofyzoidów i tetanurów, niektóre bazalne przypominają prymitywne celurozaury - szczególnie ornitomimozaury, ceratozaur ma wiele wspólnego z tetanurami, a abelizaurydy mają wiele cech wspólnych z karcharodontozaurydami i mniej z tyranozaurydami.
Ceratozaury miały wiele charakterystycznych cech czaszki (nieduże okno przedoczodołowe; kość szczękowa z krótkim wyrostkiem szczękowym oraz długim i masywnym nosowym, rozciągająca się do wewnętrznej krawędzi nozdrza; kość nosowa z przedniobocznym brzegiem nieco wypukłym bocznie, tylnobocznym brzegiem rozciągającym się do wgłębienia przedoczodołowego i bocznie wypukłym kapturem zakrywającym tylną część nozdrza zewnętrznego w bocznej powierzchni przedniej części - w okolicy tylnej krawędzi nozdrza zewnętrznego; kość łzowa z krótkim i smukłym przednim wyrostkiem - w porównaniu do brzusznego; zwrócona mocno ku przodowi kość kątowa; górne okna skroniowe łączące się nad kośćmi ciemieniowymi, tworzącymi grzebień sagitalny), kręgów (ekstremalna spneumatyzacja łuków kręgowych; parapofyzje kręgów grzbietowych wyrastające mocno na boki; kręgi krzyżowe z żebrami formującymi mniej lub bardziej ciągłą taflę w widoku wewnętrznym lub grzbietowym, obecność żeber ogonowych), kończyn przednich (duże i bardzo głębokie kości krucze; długi, niski i ukośnie skierowany grzebień deltopektoralny, kość ramienna prosta z spłaszczonymi dystalnymi kłykciami w widoku bocznym) i tylnych (zrośnięte połączenie kości łonowych u dorosłych osobników; dystalna końcówka kości łonowej z dużą, zwrócona ku tyłowi "stopą"; boczna półka dołu brevis tworząca ciągłą powierzchnię z tylną częścią półki nadpanewkowej; dystalne rozszerzenie kości łonowej z tylnym rozszerzeniem większym niż połowa długości półki łonowej; dystalny koniec kości kulszowej mocno rozrośnięty - długością ponad dwukrotnie przewyższający półki kulszowe; przednia powierzchnia dystalnej kości udowej z płytkim rowkiem prostownika; duży przeciwkrętarz kości kulszowej; duży guzek biodrowo-strzałkowy na kości strzałkowej, kość piętowa z dużą powierzchnią piszczelową) (za Carrano i Sampson, 2008 oraz Xu i in., 2009).
Duży grzebień goleniowy kości strzałkowej większości ceratozaurów sugeruje wzmocnienie mięśni odpowiadających za prostowanie kolana.
W ewolucji ceratozaurów można dostrzec wzrastającą rolę czaszki i zrastania się jej górnych kości (szczególnie zmieniła się sama natura kontaktów międzykostnych - połączenia stawały się coraz bardziej zawiłe, czasem można nawet je określić jako "wklinowanie"). Czaszki były dość wysokie, najbardziej u abelizaurydów. Ceratozaury (szczególnie abelizauroidy) cechuje też skrócona kość zębowa w porównaniu do dalszych kości żuchwy i powiększone zewnętrzne okno w żuchwie, połączone ze zmianą pozycji zetknięć kości zębowej z pozostałymi kośćmi żuchwy i rozrostem panewki między k. zębową a k. kątową górną. Jest to oznaką zmian w funkcjonowaniu stawu wewnątrzżuchwowego, być może powiązanymi ze zwiększoną ruchliwością lub większą elastycznością na rzecz utraty siły zgryzu i wytrzymałości (widoczne jest to szczególnie u abelizaurdydów, słabiej u Skorpiovenator). Prawdopodobnie powiązane ze wzrastającą rolą czaszki jest uwstecznienie kończyn przednich, w których brakuje skostniałych kości nadgarstka, a I kość śródręcza jest zredukowana i pozbawiona paliczków (choć Ceratosaurus zapewne miał paliczki, na co wskazuje powierzchnia stawowa I kości śródręcza). Przykładami karłowacenia kończyn przednich są Limusaurus i abelizaurydy. Natomiast kończyny przednie noazaurydów i Austrocheirus nie były aż tak uwstecznione, choć paliczki noazaurydów były skrócone. Noasaurus na dłoniach miał nietypowe, bardzo zakrzywione pazury.
Neoceratosauria
Bardziej zaawansowani przedstawiciele Ceratosauria należą do Neoceratosauria. Te bliżej spokrewnione z ceratozaurem należą do Ceratosauridae, których jedynym przedstawicielem jest typowy takson omawianej grupy - Ceratosaurus a drugim, potencjalnym jest Genyodectes. Oba rodzaje łączą zęby z kości szczękowych mocno ustnojęzykowo ściśnięte, przekraczające minimalną wysokość kości zębowej oraz wyraźna różnica w wysokości zębów kości przedszczękowej (i prawdopodobnie pierwszym zębem kości szczękowej) i szczękowej. Ceratozaur jest już dość masywny i pozostaje dotąd jedynym - prócz Limusaurus - bazalnym przedstawicielem Ceratosauria, którego czaszka jest znana. Mimo że jest ona bardzo charakterystyczna (choćby trzy rogi i wyjątkowo długie zęby) to zapewne prezentuje obraz najbliższy bazalnym ceratozaurom. Zasięg kladu Neoceratosauria - uzależniony od pozycji Ceratosaurus wśród innych ceratozaurów - różni się znacznie w analizach. Według niektórych (np. Ezcurra i in., 2010) ceratozaur jest najbardziej bazalnym znanym ceratozaurem a formy takie jak elafrozaur lub limuzaur są abelizauroidami (czyli nie znamy ceratozaurów niebędących neoceratozaurami), wg innych (np. Xu i in., 2009) występuje wiele nie-neoceratozaurowych ceratozaurów, natomiast kolejne (np. Sereno i Brusatte, 2008) wskazują wariant pośredni. Wyliczone niżej synapomorfie neoceratozaurów i abelizauroidów odnoszą się do tej drugiej hipotezy.
Neoceratozaury miały pleurocoel na przodzie obrotnika i otwór lub otwory pneumatyczne na jego łuku kręgowym, długie i masywne epipofyzje (epipophyses) na przednich kręgach szyjnych, przednią krawędź kości łopatkowo-kruczej gładko zakrzywioną i nieprzerwaną przez wycięcie w miejscu styku, spłaszczone dystalne kłykcie kości ramiennej, proksymalny koniec I kości śródręcza luźno przyciśnięty do II kości śródręcza, wklinowane styki kości łonowych i biodrowych oraz kulszowych i biodrowych (nie planarne, jak u innych teropodów), wyraźny, ostry grzbiet nadkłykcia przyśrodkowego kości udowej i przyśrodkowo otwarte wgłębienie na przyśrodkowej stronie proksymalnej kości strzałkowej (za Carrano i Sampson, 2008 oraz Xu i in., 2009).
Abelisauroidea i Abelisauria
Bardziej zaawansowane od ceratozaura neoceratozaury należą do Abelisauroidea Tak jak Neoceratosauria, klad ten ma zmienny zasięg w zależności od pozycji Ceratosaurus w analizie.
Berberosaurus z wczesnej jury został opisany jako abelizauroid, jednak może być on bardziej bazalnym ceratozaurem (Carrano i Sampson, 2008, Ezcurra i in., 2010) lub dilofozaurydem (Xu i in., 2009).
Abelizauroidy miały wiele charakterystycznych cech - rzeźbioną powierzchnię kości zaoczodołowych i łzowych (u abelizaurydów obejmowało to większość czaszki), zamknięty otwór podnosowy (zapewne nerwowo-naczyniowy, zlokalizowany na spojeniu kości przedszczękowej i szczękowej), pochylony przedniowewnętrznie tylny brzeg kości zaoczodołowej, brew/grzbiet lub półka/szelf wzdłuż grzbietowego brzegu kości łzowej, poziomo skierowany szew między kością potyliczną dolną a trzonem kości klinowej, przedni koniec zewnętrznego okna w żuchwie na wysokości ostatniego zęba żuchwowego, bardzo krótkie (przedniotylnie) łuki kręgowe na kręgach szyjnych (obecne u Berberosaurus), podstawę łuku kręgowego przednich kręgów ogonowych równą lub większą niż trzon (w widoku z przodu), wyrostek w kształcie skrzydła w podstawach przednich trzonów żeber szyjnych, kulistą proksymalną głowę kości ramiennej (obecne też u Limusaurus) oraz zrośnięty kontakt kości strzałkowej i wstępującego wyrostka kości skokowej (za Carrano i Sampson, 2008 oraz Xu i in., 2009).
Kolejnym wyróżnianym kladem jest Abelisauria, dzielący się na dwie grupy, które zostaną omówione osobno: Antarctometatarsus
U większości teropodów III ("środkowa") kość śródstopia jest najszersza. U kilku grup celurozaurów jest mocno zwężona (arctometatarsus), a u wielu ceratozaurów występuje sytuacja odwrotna - III kość śródstopia jest jeszcze większa i szersza, a II i IV mocno zredukowane (co nazwano antarctometatarsus). Widoczne jest to u wszystkich ceratozaurów oraz u nieopisanego, małego (<2 m) teropoda z wczesnej jury USA (Britt i in., 2010). U noazaurydów i limuzaura szczególnie mocno zwężona jest II k. śródstopia, natomiast nieco mniej u elafrozaura i prawdopodobnie także deltadroma. Dodatkowo u masiakazaura, limuzaura i deltadroma wyjątkowo zwężony jest dystalny koniec i kłykcie na IV k. śródstopia.
Taka budowa znamionuje opieranie większości ciężaru ciała na jednym palcu. Interpretuje się ją jako przystosowanie do szybszego biegu. Podobną zmianę budowy śródstopia widać u ssaków kopytnych i ptaków biegających.
Wymiary:
>0,65-0,7 m (Ligabueino) <<< ~14,8 m (Deltadromeus)
Bazalne ceratozaury były średniej wielkości - osiągały średnio 4-6 m długości. Wyjątkami są Deltadromeus (~15 m) i Limusaurus (>2 m). Potencjalnie najstarszy ceratozaur - berberozaur - osiągał ok. 5 m (porównanie z rozmiarów kości udowej z Dilophosaurus i Ceratosaurus), lecz nie był jeszcze wyrośnięty. Już w późnej jurze żyły duże ceratozaury - w Tanzanii odnaleziono fragmentaryczne szczątki przypisane do Ceratosaurus, których właściciel mógł mierzyć ponad 9 m.
Kiedy:
~170 Ma (?190-175, ?200, ?220) Ma -> 65 Ma
najstarsze:
- Limusaurus 161-156 Ma
- ?"Chuandongocoelurus" 168-161 Ma
- Spinostropheus ?168-?161 Ma
- ?Berberosaurus 190-175 Ma
najmłodsze: m. in. Carnotaurus 71-65 Ma
najmłodsze bazalne:
- Austrocheirus 72-68 Ma
Pierwszym niewątpliwym ceratozaurem jest Limusaurus z oksfordu. Obecność tak wyspecjalizowanego przedstawiciela Ceratosauria na początku późnej jury wskazuje, że jego przodek oddzielił się od innych ceratozaurów wiele milionów lat wcześniej. Wcześniejsze od niego są prawdopodobnie: chiński ceratozaur (?abelizauroid) opisany jako paratyp Chuandongocoelurus (168-161 Ma) oraz Spinostropheus (?168-?161 Ma; jednak datowanie jego szczątków jest niepewne i opiera się głównie na pozycji filogenetycznej jego oraz dwóch innych dinozaurów z formacji Tiouraren). Berberosaurus pochodzący sprzed 190-175 milionów lat może być najwcześniejszym ze znanych ceratozaurów - o ile nie jest bardziej bazalnym teropodem. Fragmentaryczne szczątki ze środkowej jury Madagaskaru (zęby i pazur; baton - 168-165 Ma; Maganuco i in., 2005; 2007) i Australii (Ozraptor - cześć piszczeli; bajos - 172-168 Ma) zostały uznane za pochodzące od ceratozaurów, prawdopodobnie abelizauroidów. W każdym razie omawiany klad musiał oddzielić się od tetanurów najpóźniej w pierwszej połowie środkowej jury (172-168 Ma), bo z tego okresu znani są już przedstawiciele tej drugiej grupy. Ceratozaury mogły powstać już w późnym triasie (zob. artykuł o tetanurach). Jeśli celofyzoidy należą do ceratozaurów, to omawiana grupa musiała powstać już 220 Ma.
Najwcześniejszymi pewnymi przedstawicielami Abelisauroidea są teropody znane z formacji Tendaguru w Afryce (~153-150 Ma), choć wiele wcześniejszych form, np. limuzaur może do nich należeć.
Do końca kredy dożyły noazaurydy, abelizaurydy oraz formy bardziej bazalne (Austrocheirus).
Gdzie:
Gondwana (Afryka, Ameryka Płd., Indie, Madagaskar, ?Australia) i Laurazja (Ameryka Płn., Azja, Europa)
Ceratozaury powstały na tyle wcześnie, że zapewne ich zasięg występowania obejmował całą Pangeę, np. późnojurajskie ceratozaurydy cechował prawdopodobnie światowy zasięg (Afryka, Ameryka Płd., Ameryka Płn., Europa). We wczesnej kredzie zasięg geograficzny ceratozaurów się skurczył - znane są z kontynentów południowych (Gondwana) oraz z Europy. Zdecydowana większość bazalnych ceratozaurów, abelizaurydów i prawdopodobnie też noazaurydów pochodzi z Afryki, więc wydaje się, że to ten kontynent był ich ojczyzną. Abelizauroidy były dominującymi pod względem ekologii i różnorodności drapieżnikami w późnej kredzie w Indiach i na Madagaskarze. W tym okresie częste były też w Ameryce Południowej, Afryce i Europie.
Dowiedz się więcej:
- o bazalnych ceratozaurach:
- o bazalnych ceratozaurach:
(Betasuchus), Ceratosaurus, Genyodectes, ("Labrosaurus" - częściwo), ?(Orthogoniosaurus)
- i abelizauroidach (trzema znakami zapytania oznaczono teropody, które mogą nie być ceratozaurami):
(?)Austrocheirus, ??(Bahariasaurus)/??Deltadromeus, ???Berberosaurus, ??"Chuandongocoelurus" (częściowo), (Coeluroides), (Compsosuchus), ??Elaphrosaurus, ??Kemkemia, ??Limusaurus, ?Ozraptor, ??Spinostropheus, ?(Tarascosaurus)

Rekonstrukcje szkieletów bazalnych ceratozaurów. Od góry: Ceratosaurus nasicornis (uznany tu za jedyny gatunek rodzaju; autor: Scott Hartman - rysunek pochodzi z tej strony), Elaphrosaurus bambergi (holotyp; autor: Jaime A. Headden - rysunek pochodzi z tej strony), Limusaurus (autor: Ville Sinkkonen - rysunek pochodzi z tej strony) i Deltadromeus agilis (holotyp; rysunek pochodzi z publikacji Sereno i in., 1996). Skala przy Elaphrosaurus i Deltadromeus = 1 m.

Rekonstrukcje czaszek bazalnych ceratozaurów. . A - Ceratosaurus (z Sampson i Witmer, 2007), B - Limusaurus (z Xu i in., 2009).

Stopa Velocisaurus unicus w widoku A - z przodu i B - z boku (zmodyfikowano z Bonaparte, 1991; rysunek pochodzi z tej strony)

Rekonstrukcja Ceratosaurus nasicornis. Autor: Julius Csotonyi. Rysunek pochodzi z tej strony.

Skalibrowany stratygraficznie kladogram z Sereno i in., 2004. Przerywaną linią oznaczono niepewne pozycje taksonów. Skala czasu za Walker i Geissman, 2009.

Skalibrowany stratygraficznie kladogram z Allain i in., 2007. Analizowani przedstawiciele Noasauridae to Masiakasaurus i Noasaurus a Abelisauridae - Abelisaurus, Aucasaurus, Carnotaurus, Ilokelesia i Majungasaurus. Skala czasu za Walker i Geissman, 2009.

Skalibrowany stratygraficznie kladogram z Sereno i Brusatte, 2008 (na podstawie Sereno i in., 2004 - z dodanym Kryptops, usuniętymi fragmentarycznymi Laevisuchus, Genusaurus i Ilokelesia oraz zmienioną nieznacznie jedną cechą). Przerywaną linią oznaczono prawdopodobną pozycję Abelisaurus (w kladogramie przedstawionym w publikacji zapewne omyłkowo zabrakło tego taksonu). Skala czasu za Walker i Geissman, 2009.

Skalibrowany stratygraficznie kladogram z Carrano i Sampson, 2008 - konsensus Adamsa. W ścisłym konsensusie Spinostropheus i Elaphrosaurus tworzyły politomię z Deltadromeus i Neoceratosauria a Carnotaurus i Ilokelesia - z pozostałymi taksonami bardziej zaawansowanymi od Ekrixinatosaurus. Skala czasu za Walker i Geissman, 2009.

Kladogram z Xu i in., 2009 - I (na podstawie Smith i in., 2007 z dodanymi 59 cechami z Carrano i Sampson, 2008 oraz dodatkowymi taksonami).

Skalibrowany stratygraficznie kladogram z Xu i in, 2009 - II, po odjęciu Elaphrosaurus, który tworzył politomię z wszystkimi ceratozaurami. Skala czasu za Walker i Geissman, 2009

Skalibrowany stratygraficznie kladogram z Canale i in., 2009. Skala czasu za Walker i Geissman, 2009.

Kladogram z Ezcurra i in., 2010 (na podstawie Canale i in., 2009 z 15 nowymi cechami i zmodyfikowanymi dwoma oraz dodanymi Austrocheirus, Berberosaurus i Limusaurus). Limusaurus przyjmował pozycję siostrzanego taksonu Masiakasaurus albo Elaphrosaurus.

Kladogram z Cau i in., 2011.
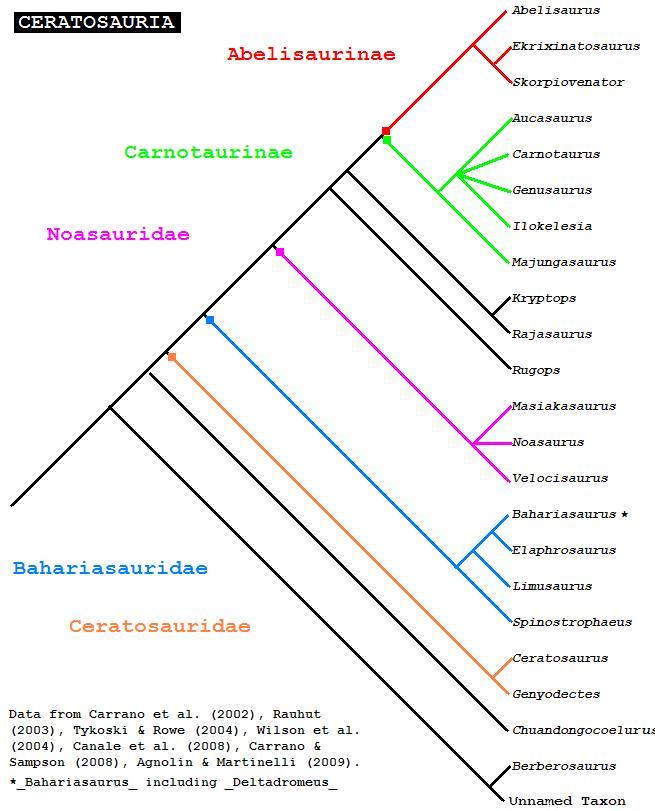
Kladogram z Cau, online. Agnolin i Martinelli, 2007 został błędnie zapisany jako 2009.
Źródła:
Publikacje naukowe:
Allain, R., Tykoski, R., Aquesbi, N., Jalil, N.-E., Monbaron, M., Russell, D. & Taquet, P. (2007) "An abelisauroid (Dinosauria: Theropoda) from the Early Jurassic of the High Atlas Mountains, and the radiation of ceratosaurs" Journal of Vertebrate Paleontology, 27, 610-624.
Canale, J.I., Scanferla, C.A., Agnolin, F.L. & Novas, F.E. (2009) "New carnivorous dinosaur from the Late Cretaceous of NW Patagonia and the evolution of abelisaurid theropods" Naturwissenschaften, 96, 409-414.
Carrano, M.T. (2007) "The appendicular skeleton of Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar" [w:] Sampson, S. D. & Krause, D. W. (ed.) "Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar" Society of Vertebrate Paleontology Memoir 8, 163–179.
Carrano, M.T., Loewen, M.A. & Sertich, J.J.W. (2011) "New Materials of Masiakasaurus knopfleri Sampson, Carrano, and Forster, 2001, and Implications for the Morphology of the Noasauridae (Theropoda: Ceratosauria)" Smithsonian Contributions to Paleobiology, 95, viii + 53.
Carrano, M.T. & Sampson, S.D. (2008) "The phylogeny of Ceratosauria (Dinosauria: Theropoda)" Journal of Systematic Palaeontology, 6, 182-236.
Cau, A., Dalla Vecchia, F.M. & Fabbri, M. (2011) "Evidence of a new carcharodontosaurid from the Upper Cretaceous of Morocco" Acta Palaeontologica Polonica, doi:10.4202/app.2011.0043
Ezcurra M.D., Agnolin F.L. & Novas F.E. (2010) "An abelisauroid dinosaur with a non-atrophied manus from the Late Cretaceous Pari Aike Formation of southern Patagonia" Zootaxa 2450, 1-25.
Madsen Jr., J. H. & Welles, S. P. (2000) "Ceratosaurus (Dinosauria, Theropoda) a revised osteology" Miscellaneous Publication Utah Geological Survey, 1-80.
Maganuco S., Cau A. & Pasini G. (2005) "First description of theropod remains from the Middle Jurassic (Bathonian) of Madagascar" Atti della Società Italiana di Scienze Naturali e del Museo Civico di Storia Naturale in Milano, 146, 165-202.
Maganuco S., Cau A., Dal Sasso C. & Pasini G. (2007) "Evidence of large theropods from the Middle Jurassic of the Mahajanga basin, NW Madagascar, with implications for the ceratosaurian pedal ungual evolution" Atti della Società Italiana di Scienze Naturali e del Museo Civico di Storia Naturale in Milano, 148, 261-271.
Novas F.E., Ezcurra M.D. & Agnolin F.L. (2006) "Humerus of a basal abelisauroid theropod from the Late Cretaceous of Patagonia" Revista del Museo Argentino de Ciencias Naturales, nueva serie 8, 63-68.
Rauhut, O.W.M. (2003) "The interrelationships and evolution of basal theropod dinosaurs" Special Papers in Palaeontology, 69, 1-213.
Rauhut, O.W.M. (2004) "Provenance and anatomy of Genyodectes serus, a large-toothed ceratosaur (Dinosauria: Theropoda) from Patagonia" Journal of Vertebrate Paleontology, 24, 894-902.
Rauhut, O.W.M. (2011) "Theropod Dinosaurs from the Late Jurassic of Tendaguru (Tanzania)" Special Papers in Palaeontology, 86, 195-239. doi: 10.1111/j.1475-4983.2011.01084.x
Rauhut, O.W.M. & López-Arbarello, A (2009)" Considerations on the age of the Tiouaren Formation (Iullemmeden Basin, Niger, Africa): Implications for Gondwanan Mesozoic terrestrial vertebrate faunas" Palaeogeography, Palaeoclimatology, Palaeoecology, 271, 259–267. doi:10.1016/j.palaeo.2008.10.019
Soto, M. & Perea, D. (2008) "A ceratosaurid (Dinosauria, Theropoda) from the Late Jurassic-Early Cretaceous of Uruguay" Journal of Vertebrate Paleontology, 28(2), 439–444. doi:10.1671/0272-4634(2008)28[439:ACDTFT]2.0.CO;2
Sereno, P.C. & Brusatte, S.L. (2008) "Basal abelisaurid and carcharodontosaurid theropods from the Lower Cretaceous Elrhaz Formation of Niger" Acta Palaeontologica Polonica, 53, 15-46.
Sereno, P.C., Wilson, J.A. & Conrad, J.L. (2004) "New dinosaurs link landmasses in the Mid-Cretaceous" Proceedings of the Royal Society of London B: Biological Sciences, 271, 1325-1330.
Smith N.D., Makovicky P.J., Pol D., Hammer W.R. & Currie P.J. (2007) "Osteology of Cryolophosaurus ellioti (Dinosauria: Theropoda) from the Early Jurassic of Antarctica and implications for early theropod evolution" Zoological Journal of the Linnean Society 151, 377-421
Tykoski, R.S. & Rowe, T. (2004) "Ceratosauria" [w:] Weishampel, D.B., Dodson, P. & Osmólska, H. "The Dinosauria" Berkeley and Los Angeles: University of California Press, 151-168.
Wilson, J.A., Sereno, P.C., Srivastava, S., Bhatt, D.K., Khosla, A. & Sahni, A. (2003) "A new abelisaurid (Dinosauria, Theropoda) from the Lameta Formation (Cretaceous, Maastrichtian) of India" Contributions from the Museum of Paleontology, the University of Michigan, 31, 1-42.
Xu, X., Clark, J.M., Mo J., Choiniere, J., Forster, C.A., Erickson, G.M., Hone, D.W.E., Sullivan, C., Eberth, D.A., Nesbitt, S., Zhao, Q., Hernandez, R., Jia, C., Han, F. & Guo, Y. (2009) "A Jurassic ceratosaur from China helps clarify avian digital homologies" Nature, 459, 940-944.
Inne:
Britt, B., Chure, D., Engelmann, G., Sheetz, R. & Hansen, R. (2010) "Multi-taxic theropod bonebeds in an interdunal setting of the Early Jurassic eolian nugget sandstone, Utah" Society of Vertebrate Paleontology Seventienth Anniversary Meeting Program and Abstracts Book, 65A [abstrakt]
http://www.forum.dinozaury.com/posting. ... =6&p=33960