


Jesteś w: Artykuły o teropodach
Tetanury (Tetanurae) bazalne
Autor: Maciej Ziegler, korekta: Tomasz Sokołowski
Ostatnia aktualizacja: 1.12.2011 r.
Nomenklatura:
- Tetanurae (Gauthier, 1986)
>Passer domesticus ~ Ceratosaurus nasicornis (Holtz, Molnar i Currie, 2004; zmodyfikowano z Padian, Hutchinson i Holtz, 1999; zmodyfikowano z Gauthier, 1986) - Megalosauroidea (Huxley, 1869 sensu Nopcsa, 1928)
>Megalosaurus bucklandii ~ Allosaurus fragilis v Passer domesticus (Benson, 2010)
Megalozauroidy często nazywa się spinozauroidami, które definiuje się dwojako: jako najszerszy klad zawierający spinozaura, ale nie wróbla (Holtz i in., 2004) lub najwęższy klad zawierający wszystkich potomków najmłodszego wspólnego przodka spinozaura i torwozaura (Sereno, 1998 - lecz wtedy spinozauroidami mogą być niemal wszystkie tetanury - zob. Sadleir i in., 2008). Tradycyjnie uważa się, że zasięgi tak definiowanych kladów niemal pokrywają się (tzn. nie ma spinozauroida w znaczeniu szerszym, który nie byłby też spinozauroidem w znaczeniu węższym). Jednak wg nowszych badań (Smith i in., 2007 i 2008; oraz szczególnie Benson, 2010 A), wiele taksonów jest spinozauroidami sensu Holtz i in., 2004 (=Megalosauroidea sensu Benson, 2010), ale już nie Sereno, 1998. Dlatego w dalszej części artykułu będziemy używać nazwy Spinosauroidea na określenie kladu o węższym zasięgu (takim, jaki nadał mu najpierw Sereno), pamiętając jednak, że w myśl starej klasyfikacji linneuszowskiej nieprawidłowe jest zaliczenie jednej nadrodziny do drugiej (dla Phylocode nie stanowi to oczywiście problemu). Taksonem kluczowym dla przedstawionej tutaj nomenklatury jest Megalosaurus bucklandii, którego jedyny pewny materiał (fragment żuchwy - zob. jego opis) wykazuje jedną unikalną cechę spinozauroidów (a przynajmniej megalozauroidów bardziej zaawansowanych od Monolophosaurus) - szeroki z przodu rowek przyzębowy na kości zębowej (u pozostałych teropodów jest on wąski na całej długości) - a spośród nich najbardziej przypomina budową Torvosaurus. Ponieważ takson Megalosauroidea (Huxley, 1869 sensu Nopcsa, 1928) został utworzony wcześniej niż Spinosauroidea (Stromer, 1915 sensu Olshevsky, 1991), to jako nadrodzina ma pierwszeństwo - analogicznie rodziny Megalosauridae (Huxley, 1869) i Torvosauridae (Jensen, 1985). Torvosauridae została zdefiniowane wcześniej (Sereno, 1998), więc w myśl Phylocode to ona - jako nazwa kladu - ma pierwszeństwo. Podobnie Torvosaurinae (Jensen 1985 sensu Allain, 2002) i Megalosaurinae (Huxley, 1869 sensu Nopcsa, 1928)- oba klady zostały po raz pierwszy zdefiniowane przez Allaina (2002), lecz ten pierwszy (który zapewne jest szerszy - zob. Benson i in., 2010) akapit (!) wcześniej. - Spinosauroidea (Stromer, 1915 sensu Olshevsky, 1991)
<Spinosaurus aegyptiacus & Torvosaurus tanneri (zmodyfikowano z Sereno, 1998) - Avifilopluma (Gauthier i de Queiroz, 2001)
> pióra (=nitkowate struktury wyrastające z brodawek skórnych) (Vultur gryphus)
Klad o niepewnym zasięgu. Może pokrywać się z Avetheropoda, jeśli Concavenator rzeczywiście miał pióra albo być ograniczony jedynie do Coelurosauria. Wg innego poglądu należą do niego wszystkie Ornithodira sensu Benton (2004): Pterosauria i Dinosauromorpha. Niepublikowane dane wskazują, że do Avifilopluma należą praktycznie wszystkie tetanury. - Avetheropoda (Paul, 1988) / Neotetanurae (Sereno, Wilson, Larsson, Duthell i Sues, 1994
<Allosaurus fragilis & Passer domesticus (Holtz, Molnar i Currie, 2004; zmodyfikowano z Padian, Hutchinson i Holtz, 1999; zmodyfikowano z Currie i Padian, 1997)
Dla tego kladu używa się często i być może poprawnie nazwy Neotetanurae (Sereno, Wilson, Larsson, Duthell i Sues, 1994), zdefiniowanej w 1998 r. przez Sereno. Currie i Padian (1997) błędnie uznali, że nazwę Avetheropoda zdefiniował Holtz (1994) i przytoczyli przypisaną jemu definicję. Trudno stwierdzić, czy to, co napisali Currie i Padian należy uznać za ich własną definicję. Jeśli nie, to prawidłową nazwą jest Neotetanurae. W artykule używa się nazwy Avetheropoda, gdyż jest starsza, ma dwie potencjalnie ważne definicje sprzed 1998 r. i jest popularniejsza (na dzień 22.10.2010 r. Google Scholar dają ok. 78 wyników, a dla Neotetanurae - ok. 48).
Charakterystyka:
Tetanury to bardzo liczna i zróżnicowana grupa teropodów. Zamieszkiwały (i nadal zamieszkują) rozmaite środowiska, wcielając się w przeróżne role w ekosystemach. Jednak nie od zawsze tak było - pierwsze tetanury były średnimi i dużymi, typowo lądowymi, niezbyt wyspecjalizowanymi drapieżnikami. Ich nazwa - "sztywne ogony" została nadana przez Jacquesa Gauthiera, jednego z pionierów kladystyki w paleontologii kręgowców, który w artykule z 1986 roku wprowadził nową metodę do "dinozaurologii". Nazwa ta odnosi się do tylnych kręgów ogonowych, które zazębiały się - co prawdopodobnie działało jako stabilizator podczas biegu i pozwalało na szybsze zwroty.
Bazalne tetanury miały raczej mocne czaszki z ostrymi zębami skupionymi z jej przodu (w widoku bocznym nie wyrastały za linię oczodołu) i dobrze rozwiniętymi oknami w kościach szczękowych.
Szyje oraz kończyny przednie wczesnych tetanurów były silne, co w połączeniu z uzbrojoną w sztyletowate zęby paszczą czyniło je niebezpiecznymi drapieżnikami.
Wśród bazalnych tetanurów doszło do gwałtownej ewolucji budowy kończyn przednich - szczególnie w dłoni (Dececchi i Larsson, 2009), które później zaowocowały pojawieniem się form latających - Avialae. Cechuje je duża dystalna kość nadgarstka, pokrywająca I i część II kości śródręcza, I kość śródręcza przyciśnięta do proksymalnej połowy II i proksymalny koniec III kość śródręcza w kształcie klepsydry - przyśrodkowa i/lub boczna powierzchnia/e grzbietowowewnętrznie wklęsłe. Nadgarstek stał się bardziej mobilny.
Tetanury cechują się niskim, zgrubiałym środkowym grzbietem na bocznej stronie kości biodrowej i szeroko rozmieszczonymi prezygapofyzjami (prezygapophyses) kręgów szyjnych. Inne synapomorfie tego kladu to (za Smith i in., 2007; Rauhut, 2003): pneumatyczne otwory powiązane z wewnętrzną tętnicą szyjną, "prętowaty" wyrostek kolczysty obrotnika, masywne wewnętrzne kile na przednich kręgach grzbietowych, skierowana poziomo głowa kości udowej, kłykieć strzałkowy na piszczeli mocno oddalony od strzałkowego grzebienia goleniowego (obecne też u Limusaurus), ulokowany dystalnie grzebień strzałkowy na piszczeli, III kość śródstopia z tylnym konturem w kształcie klepsydry, IV kość śródstopia z dobrze rozwiniętym tylnośrodkowym kołnierzem, redukcja tylnych pleuroceli w kręgach szyjnych, mniejszy krętarz ulokowany proksymalnie - niżej niż większy.
Tetanury - jak wynika ze zdecydowanej większości badań - dzielą się na dwie duże grupy - megalozauroidy (Megalosauroidea, zwane także spinozauroidami - Spinosauroidea) i aweteropody (Avetheropoda - inaczej neotetanury - Neotetanurae).
1. Megalosauroidea i Spinosauroidea
Megalosauroidea to grupa mniej liczna i zróżnicowana niż aweteropody. Cechami charakterystycznymi są: U-kształtny brzuszny wyrostek kości zaoczodołowej, proksymalna powierzchnia stawowa szewronów bez wyraźnych płaszczyzn przednich i tylnych oraz okno w kości szczękowej w formie niemal kulistego dołu (synapomorfie za Benson, 2010 A).
Do Megalosauroidea należą, prócz prawdopodobnie kilku bazalnych kladów (zob. Benson, 2010 A), także spinozauroidy. Cechują się one dość krótkimi i mocnymi kończynami przednimi oraz wydłużonymi czaszkami. Wszystkie miały też nozdrza ulokowane za szeregiem zębów przedszczękowych, osłonę kości potylicznej dolnej (basioccipital apron) z szerokim (ok. 2/3 szerokości kłykcia potylicznego) obniżeniem, rowek przyzębowy na kości zębowej szeroki z przodu i wąski z tyłu i nie miały proksymalnej wklęsłości na środkowej powierzchni kości strzałkowej. Wśród nich wyróżnia się omówione osobno:
- Spinosauridae - to grupa dużych, wyspecjalizowanych, wyraźnie odrębnych i niewątpliwie tworzących klad tetanurów z kredy.
- Megalosauridae (Torvosauridae) - grupa jurajskich, niewyspecjalizowanych, średnich-dużych tetanurów, które mogą nie tworzyć kladu.
U aweteropodów pojawiło się kilka zmniejszających ciężar przystosowań - dodatkowe okno w czaszce (w kości szczękowej) i bardzo złożone komory powietrzne w kręgach (np. pleurocele w obrotniku; obecne też u abelizaurydów i zauropodów). Inne cechy charakterystyczne (synapomorfie za Smith i in., 2007 i Benson, 2010 A) to m. in. krótka przednia gałąź kości łzowej i redukcja szerokości wewnętrznej gałęzi tej kości, wyraźny nadkłykieć łokciowy na kości ramiennej oraz klinowaty kształt i ściśnięta powierzchnia podeszwowa III kości śródstopia.
Avetheropoda dzielą się na:
- Carnosauria
- Coelurosauria
omówione w osobnych działach.
Prawdopodobnie wszyscy przedstawiciele Avetheropoda byli częściowo pokryci pra-piórami, choć nie jest to pewne (zob. opis Concavenator), bo mogła być to cecha jedynie celurozaurów, i to nie wszystkich (na pewno kladu Tyrannoraptora+Compsognathidae, lecz niektóre celurozaury, np. Tugulusaurus, znajdują się prawdopodobnie poza tym taksonem). Jednakże może to być cecha obszerniejszego kladu - nie jest znane pokrycie ciała bardziej bazalnych tetanurów, np. megalozauroidów, a nawet gdyby odnaleziono odciski łusek, nie znaczy to, że piór nie było. Niepublikowane dane wskazują, że pierzaste: Juravenator i jego nienazwany krewniak są bazalnymi megalozauroidami (zob. komentarz na DML). Jeśli jest tak rzeczywiście, oznacza to, że praktycznie wszystkie obecnie znane tetanury miały prapióra. Zheng i in. (2009) wskazują, że pierwotnie "opierzone" były wszystkie dinozaury (zob. opis Tianyulong). Pewne nieopublikowane badania Longricha z 2002 roku wskazują, że Sinosauropteryx (uważany za kompsognatyda) jest najbardziej bazalnym celurozaurem albo allozaurydem, co wskazywałoby na to, że upierzenie jest cechą aweteropodów, do których jednak nie należały sinraptorydy.
Redukcja liczby palców u tetanurów
Palce i kości śródręcza kręgowców oznacza się rzymskimi liczbami I od V, gdzie I to kciuk, a V nasz mały palec. Dłonie teropodów miały utracony lub znacznie zredukowany palec V. Wśród tetanurów doszło do zmniejszenia liczby palców, co uczyniło ich kończyny trójpalczastymi. Znanych jest jednak kilka wyjątków z zachowaną IV kością śródręcza - bazalnych celurozaurów (Coelurus, Guanlong, Tanycolagreus), karnozaurów (Megaraptor, Sinraptor i wg Mortimera, online także Erectopus) i bardziej bazalnych tetanurów (Szechuanosaurus? zigongensis, Xuanhanosaurus). Kości dłoni tetanurów tradycyjnie brano za I-II-III (i czasem IV), co stoi w sprzeczności z badaniami embrionów ptaków, które - choć zostało to podważone - mają palce II-III-IV. Xu i współpracownicy (2009) uznali ceratozaura Limusaurus za dowód kopalny hipotezy traktującej o tym, że palce tetanurów to w rzeczywistości II-III-IV, co jest jednak nieprzekonujące (zob. Cau, online i Mortimer, online).
Wymiary:
[5 cm (Mellisuga helenae)], ~43 cm (Parvicursor) <<< 16-18 m (Spinosaurus)
Tetanury to najbardziej zróżnicowana grupa teropodów - także pod względem rozmiarów. Należą do nich zarówno najmniejsze (nawet nie licząc ptaków), jak i największe teropody. Bazalne (niecelurozaurowe) tetanury były przeważnie dość duże - miały ok. 4-8 m długości, a wiele z nich osiągnęło w toku ewolucji znacznie większe rozmiary - wiele spinozaurydów, megalozaurydów i karnozaurów miało po kilkanaście metrów długości oraz kilka ton wagi; z tytonu (151-146 Ma) Tanzanii pochodzą kości kończyn megalozauroida większego niż Torvosaurus. Niewielkim wyjątkiem mógł być Chuandongocoelurus mierzący ok. 2 m i ważący 13-14 kg, lecz może chodzić o młodego osobnika.
Znaczna redukcja rozmiarów nastąpiła u celurozaurów - ale o tym w rozdziale im poświęconym.
Kiedy:
~170 Ma (?190-175, ?200, ?220) Ma -> dziś
najstarsze:
Shidaisaurus - 176-167 Ma
Magnosaurus - 172-168 Ma
?Kayentavenator - 193-187 Ma
??Cryolophosaurus - 203-177 Ma (bliżej 203)
najmłodsze bazalne: Marshosaurus ~ 153-150 Ma
Najstarszymi niewątpliwymi tetanurami są środkowojurajskie Magnosaurus i Shidaisaurus, jednak wiek tego drugiego jest wielce niepewny. Wczesnojurajski Kayentavenator (późny synemur-wczesny pliensbach, USA) miałby być najwcześniejszym z nich, lecz w środowisku naukowym trwają spory o ważność tego taksonu i przynależność do omawianego kladu (zob. Cau, online). Cryolophosaurus, długo uważany za tetanura, okazał się bardziej bazalnym teropodem (Smith i in., 2007; Xu i in., 2009; Nesbitt i in., 2009), lecz niektóre z nowszych analiz - wykonana przez Choiniere i in. (2010) oraz druga z analiz Xu i in. (2009) - wykazała znów tetanurową naturę kriolofozaura.
Wcześniej za tetanury uważano m.in. Zupaysaurus z noryku-?retyku (228-204, -?201 Ma) Argentyny, "Zanclodon" cambrensis z retyku (204-199 Ma) Anglii, "Merosaurus" z hettangu-synemuru (200-190 Ma) Anglii (który może być jednak tetanurem, zob. Cau, online), teropoda z toarku (183-175 Ma) Maroko (który według nieopublikowanych analiz - Allain i Bailleul, 2009 - jest najbardziej bazalnym tetanurem), kręgu "Streptospondylus" cuvieri z toarku (183-175 Ma) Anglii (może być tetanurem albo zauropodem) i "Saltriosaurus" z synemuru (196-190 Ma) Włoch. Nieoficjalnie podawano, że Smok (205-200 Ma) także jest tetanurem (zob. temat na forum), lecz obecnie uważa się, że chodzi na pewno o archozaura, lecz nie jest pewne, czy o rauizucha (z "krokodylej" linii archozaurów), czy teropoda. Jeśli celofyzoidy są ceratozaurami, to tetanury musiały już istnieć wtedy, kiedy najstarsze celofyzoidy, czyli ok. 220 Ma. Obecność stosunkowo zaawansowanego Magnosaurus we wczesnym bajosie i niewiele późniejszych tyranozauroidów w batonie wskazuje na to, że radiacja tetanurów miała miejsce dużo wcześniej - prawdopodobnie już we wczesnej jurze (zob. też artykuł o ceratozaurach).
Okresem świetlności nieaweteropodowych tetanurów była środkowa jura - były wtedy czołowymi dużymi drapieżnikami. W późnej jurze zostały w większości wyparte przez karnozaury, a te, które przetrwały, były rzadkie.
Do niedawna sądzono, że do końca mezozoiku nie dotrwały żadne niecelurozaurowe tetanury. Prawdopodobnie najwcześniej, bo zapewne jeszcze w jurze, wyginęły formy najprymitywniejsze oraz megalozaurydy. Ostatni z nich - Torvosaurus - dotrwał do późnego tytonu (151-145 Ma), natomiast z nieco wcześniejszego okresu (późny kimeryd, 153-150 Ma) znany jest ostatni z bardziej bazalnych megalozauroidów - Marshosaurus (choć jego pozycja filogenetyczna jest bardzo niepewna - może być przedstawicielem innej grupy albo nawet chimerą). Z osadów późnokredowych znane są jednak fragmentaryczne szczątki pochodzące - jak się wydaje - od megalozaurydów (zob. artykuł o nich). Dystalny fragment piszczeli pochodzący od bardzo dużego teropoda z santonu (86-83 Ma) Antarktydy wykazuje podobieństwo do Piatnitzkysaurus (Molnar i in., 1996; Case i in., 2007). Spinozaurydy radziły sobie nieźle - na nie kolej przyszła dopiero w późnej kredzie (zob. artykuł o nich). Z wczesnego mastrychtu (72-68 Ma) znany jest (Orkoraptor) - przedstawiciel megaraptorów (zob. artykuł o karnozaurach).
Gdzie:
Nie wiemy, gdzie pojawiły się tetanury. Prawdopodobnie powstały dość wcześnie a ich zasięg geograficzny obejmował całą Pangeę. W większości nieopisane taksony, które mogą być najstarszymi lub najbardziej bazalnymi tetanurami znane są z Afryki, Azji i szczególnie z Europy. W środkowej jurze tetanury prawdopodobnie cechowały się przynajmniej częściowym endemizmem, choć może to wynikać z niekompletności zapisu kopalnego i nieznanej lub niepewnej pozycji filogenetycznej wielu taksonów, głównie azjatyckich. Siostrzane taksony Condorraptor i Piatnitzkysaurus znane są z Ameryki Południowej. Szczególnie wiele tetanurów z tego okresu znanych jest z Europy - wszystkie środkowojurajskie megalozaurydy pochodzą właśnie stamtąd.
Późniejsze tetanury żyły niemal wszędzie (choć np. w późnojurajskich osadach z Ameryki Płd. dotychczas ich nie znaleziono). Stan ten trwa do dziś.
Dowiedz się więcej:
- o tetanurach o niejasnej klasyfikacji:
Cruxicheiros, ??(Dandakosaurus), (Kaijiangosaurus), (Kakuru), Kayentavenator, "Mifunesaurus", (Prodeinodon), "Scrotum", Sigilmassasaurus, "Szechuanoraptor"
- megalozauroidach:
??Chuandongocoelurus, ??Condorraptor, ??Marshosaurus, ??Monolophosaurus, ??Piatnitzkysaurus, ??Xuanhanosaurus
- i aweteropodach o niejasnej klasyfikacji:
(?)Gasosaurus, ?(Iliosuchus), Shanyangosaurus

Szkielet Piatnitzkysaurus floresi. Na szaro - znaleziony materiał, na biało - zrekonstruowane kości. Autor: Jorge Gonzalez. Rysunek pochodzi z publikacji Escapa i in. (2008).

Rekonstrukcja Monolophosaurus jiangi. Autor: Michael Skrepnick. Rysunek pochodzi z tej strony.

Skalibrowany stratygraficznie kladogram z Charig i Milner, 1997 (na podstawie Holtz, 1994 ze zmianami i dodanym Baryonyx). Skala czasu za Walker i Geissman, 2009.

Skalibrowany stratygraficznie kladogram z Chure, 2000/nieopublikowane. Przerywaną linią zaznaczono niepewną pozycję Neovenator. Skala czasu za Walker i Geissman, 2009.

Skalibrowany stratygraficznie kladogram z Holtz, 2000. Skala czasu za Walker i Geissman, 2009.

Kladogram z Allain, 2002 (tylko cechy czaszki). Zasięg czasowy spinozaurydów rozszerzono o nieanalizowanego Spinosaurus (gdyż Allain używa nazw Spinosauridae i Spinosauroidea) a celurozaurów o nieanalizowanego Proceratosaurus (gdyż wszystkie badania wskazują, że to celurozaur).

Kladogram z Rauhut, 2003. Z analizy usunięto ostatecznie Poekilopleuron (Carnosauria incertae sedis, w matrycy zawierającej wszystkie taksony Tetanurae incertae sedis), Shaochilong (Coelurosauria incertae sedis, w matrycy zawierającej wszystkie taksony Tetanurae incertae sedis), Siamotyrannus (Allosauroidea incertae sedis) i Xuanhanosaurus (Tetanurae incertae sedis).
Wiele taksonów zakodowano tak, aby zawierały kilka rodzajów:
- Magnosaurus zawiera Eustreptospondylus
- Baryonychidae (synonim Spinosauridae): Angaturama, Baryonyx, Irritator, Spinosaurus, Suchomimus
- Carcharodontosauridae: Acrocanthosaurus, Carcharodontosaurus, Giganotosaurus).
- Sinraptoridae: Yangchuanosaurus magnus, Y. shangyouensis, Sinraptor dongi, S. hepingensis
W ścisłym konsensusie wśród Tetanurae wyróżniono jedynie następujące klady, tworzące politomię z pozostałymi taksonami: Maniraptoriformes, Allosauroidea (zawierający Siamotyrannus) i Spinosauroidea.

Kladogram z Holtz i in., 2004. Szechuanoraptor to Szechuanosaurus zigongensis, "P." valesdunensis to Dubreuillosaurus valesdunensis, do Sinraptoridae należą Sinraptor i Yangchuanosaurus.

Skalibrowany stratygraficznie kladogram z Yates, 2005 (na podstawie Rauhut, 2003 z połączeniem celurozaurów bardziej zaawansowanych od Proceratosaurus w jedną jednostkę, wyłączeniem Acrocanthosaurus z Carcharodontosauridae, rozbiciem Megapnosaurus na gatunki, dodaniem Dracovenator, Masiakasaurus, Dubreuillosaurus, Tugulusaurus i Zupaysaurus oraz 28 nowych cech i rewizją 5). Skala czasu za Walker i Geissman, 2009.

Kladogram z Xu i in., 2006 - Adams consensus (na podstawie Rauhut, 2003 z dodanymi Guanlong i Dilong oraz odjętym Deltadromeus). W ścisłym konsensusie wszystkie taksony należące do siostrzanego kladu celurozaurów (szerokie Carnosauria), z wyjątkiem kladu (Torvosaurus (Chilantaisaurus + Baryonychidae)) tworzyły politomię. S-zigongensis to Szechuanosaurus zigongensis.

Kladogram ze Smith i in., 2007.

Skalibrowany stratygraficznie kladogram z Sadleir i in., 2008 (na podstawie Holtz, 2000 ze zmienionym Eustreptospondylus). Skala czasu za Walker i Geissman, 2009.

Kladogram z Xu i in., 2009 - I (na podstawie Smith i in., 2007 z dodanymi 59 cechami z Carrano i Sampson, 2008 oraz dodatkowymi taksonami). NMV P186076 to australijski megaraptor opisany przez Smitha i in. (2008).

Kladogram z Xu i in., 2009 - II.

Kladogram z Choiniere i in., 2010B (na podstawie Choiniere, nieopublikowane).

Kladogram z Rauhut, Milner i Moore-Fay, 2010 (na podstawie matrycy Rauhuta, 2003; z wieloma zmianami mającymi na celu zbadanie pozycji filogenetycznej z Proceratosaurus). Takson Spinosauroidea został zakodowany jako jeden OTU (operational taxonomic unit).

Skalibrowany stratygraficznie kladogram z Benson, 2010 A. W ścisłym konsensusie uwidoczniły się jedynie cztery klady: Coelurosauria (zawierający Chilantaisaurus, Compsognathus, Guanlong i Tanycolagreus), Spinosauridae, Condorraptor + Piatnitzkysaurus oraz Chuandongocoelurus + Monolophosaurus. Po odjęciu Eocarcharia, Magnosaurus, Megaraptor, Piveteausaurus, Poekilopleuron i Streptospondylus uzyskano widoczny wynik. Przerywanymi liniami zaznaczono niepewne pozycje ww. taksonów. Skala czasu za Walker i Geissman, 2009.

Skalibrowany stratygraficznie kladogram z Benson i in., 2010 (na podstawie Benson, 2010 A - z wieloma zmianami - dodaniem Aerosteon, Australovenator, Orkoraptor i Shaochilong; zmianą Chilantaisaurus, Megaraptor i Neovenator; dodaniem cech różnicujących ww. tetanury oraz wzmacniających monofiletyczność ceratozaurów na podstawie Carrano i Sampsona, 2008). W ścisłym konsensusie wyżej widoczne megalozaurydy (oraz Piveteausaurus) tworzyły politomię ze Spinosauridae, tworząc Spinosauroidea sensu Sereno, 1998 (<Spinosaurus aegyptiacus & Torvosaurus tanneri). Politomię u podstawy Tetanurae tworzyły, prócz Avetheropoda, Megalosauroidea oraz kladu tworzonego przez Condorraptor, Piatnitzkysaurus, Marshosaurus i Xuanhanosaurus także Chuandongocoelurus i Monolophosaurus (tego pierwszego - jako problematycznego - ostatecznie usunięto z analizy). Taksony tworzące klad bardziej zaawansowanych niż Neovenator neowenatorydów tworzyły politomię. Po usunięciu z analizy problematycznego Orkoraptor uzyskano widoczny wynik. Przerywaną linią zaznaczono niepewną pozycję tego taksonu. Skala czasu za Walker i Geissman, 2009.

Kladogram z Ortega i in., 2010 - konsensus z zasadą większości [majority rule] (na podstawie Benson i in, 2010 z dodaniem Concavenator i usunięciem fragmentarycznych Chilantaisaurus, Piveteausaurus, Poekilopleuron, Streptospondylus, Xuanhanosaurus oraz Orkoraptor). W ścisłym konsensusie politomię u podstawy Tetanurae tworzyły Spinosauroidea, Avetheropoda, Chuandongocoelurus, Monolophosaurus i klad Marshosaurus(Condorraptor+Piatnitzkysaurus). Także megalozaurydy tworzyły politomię z jednym wyróżnionym kladem: (Megalosaurus+Torvosaurus). U podstawy Carcharodontosauria politomię tworzyły Neovenatoridae, Carcharodontosauridae, Eocarcharia i Concavenator a jedynym kladem w Carcharodontosauridae był Carcharodontosaurinae (pozostałe tworzyły politomię).

Kladogram z Cau i in., 2011.
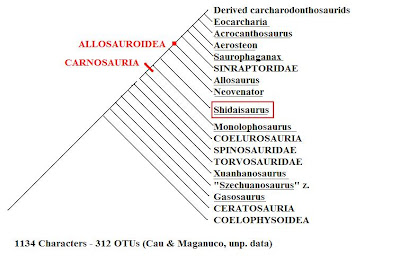
Kladogram z Cau i Maganuco (nieopublikowane - online).
Źródła
Publikacje naukowe:
Agnolin, F.L., Ezcurra, M.D., Pais, D.F. & Salisbury, S.W. (2010) "A reappraisal of the Cretaceous non-avian dinosaur faunas from Australia and New Zealand: evidence for their Gondwanan affinities" Journal of Systematic Palaeontology 8, 257-300.
Allain, R. (2002) "Discovery of megalosaur (Dinosauria, Theropoda) in the middle Bathonian of Normandy (France) and its implications for the phylogeny of basal Tetanurae" Journal of Vertebrate Paleontology 22, 548-563.
Benson, R.B.J. (2009) "An assessment of variability in theropod dinosaur remains from the Bathonian (Middle Jurassic) of Stonesfield and New Park Quarry, UK and taxonomic implications for Megalosaurus bucklandii and Iliosuchus incognitus" Palaeontology 52, 857-877.
Benson, R.B.J. (2010 A) "A description of Megalosaurus bucklandii (Dinosauria: Theropoda) from the Bathonian of the UK and the relationships of Middle Jurassic theropods" Zoological Journal of the Linnean Society, 158, 882-935.
Benson, R.B.J. (2010) "The osteology of Magnosaurus nethercombensis (Dinosauria, Theropoda) from the Bajocian (Middle Jurassic) of the United Kingdom and a re-examination of the oldest records of tetanurans" Journal of Systematic Palaeontology 8, 131-146.
Benson, R.B.J. & Xu, X. (2008) "The anatomy and systematic position of the theropod dinosaur Chilantaisaurus tashuikouensis Hu, 1964 from the Early Cretaceous of Alanshan, People's Republic of China" Geological Magazine, 145,778-789.
Benson, R.B.J. & Radley, J.D. (2010) "A new large-bodied theropod dinosaur from the Middle Jurassic of Warwickshire, United Kingdom" Acta Palaeontologica Polonica, 55, 35-42.
Benson, R.B.J., Carrano, M.T. & Brusatte, S.L. (2010) "A new clade of archaic large-bodied predatory dinosaurs (Theropoda: Allosauroidea) that survived to the latest Mesozoic" Naturwissenschaften, 97, 71-78.
Barrett, P.M., Kear, B.P. & Benson, R.B.J.(2010) "Opalized archosaur remains from the Bulldog Shale (Aptian: Lower Cretaceous) of South Australia" Alcheringa: An Australasian Journal of Palaeontology, 34, 293-301.
Brusatte, S. L., Nesbitt, S. J., Irmis, R. B., Butler, R. J., Benton, M. J. & Norell, M. A. (2010A) "The origin and early radiation of dinosaurs" Earth-Science Reviews, 101, 68-100.
Brusatte, S. L., Benson, R.B.J. & Xu, X. (2010B) "The evolution of large-bodied theropod dinosaurs during the Mesozoic in Asia" Journal of Iberian Geology, 36(2), 275-296.
Case, J.A., Martin, J.E. & Reguero, M. (2007) "A dromaeosaur from the Maastrichtian of James Ross Island and the Late Cretaceous Antarctic dinosaur fauna" [w:] "Antarctica: A Keystone in a Changing World" Online Proceedings of the 10th ISAES X Cooper, A.K. & Raymond, C.R. et al. [ed.], USGS Open-File Report 2007-1047, Short Research Paper 083.
Cau, A., Dalla Vecchia, F.M. & Fabbri, M. (2011) "Evidence of a new carcharodontosaurid from the Upper Cretaceous of Morocco" Acta Palaeontologica Polonica, doi:10.4202/app.2011.0043
Charig, A.J. & Milner, A.C. (1997) "Baryonyx walkeri, a fish-eating dinosaur from the Wealden of Surrey" Bulletin of the Natural History Museum of London, 53, 11–70.
Choiniere, J.N., Xu, X., Clark, J.M., Forster, C.A., Guo, Y. & Han, F. (2010A) "A Basal Alvarezsauroid Theropod from the Early Late Jurassic of Xinjiang, China" Science, 327, 571-57
Choiniere, J.N., Clark, J.M., Forster, C.A. & Xu, X. (2010B) "A basal coelurosaur (Dinosauria: Theropoda) from the Late Jurassic (Oxfordian) of the Shishugou Formation in Wucaiwan, People's Republic of China" Journal of Vertebrate Paleontology, 30(6), 1773-1796. doi: 10.1080/02724634.2010.520779
Gay, R. (2010) "Kayentavenator elysiae, a new tetanuran from the early Jurassic of Arizona" [w:] "Notes on Early Mesozoic Theropods" wyd. Lulu, 27-43.
Holtz, T.R. (2000) "A new phylogeny of the carnivorous dinosaurs" Gaia, 15, 5-61.
Holtz, T.R., Molnar, R.E. & Currie, P.J. (2004) "Basal Tetanurae" [w:] Weishampel, D.B., Dodson, P. & Osmólska, H. "The Dinosauria" Berkeley and Los Angeles: University of California Press, 71-110.
Lim, J.-D., Martin, L.D. & Baek, K.S. (2002) "The first megalosaurid tooth from South Korea" Current Science, 82,326-328.
Niedźwiedzki, G., Sulej, T. & Dzik, J. (2011) "A large predatory archosaur from the Late Triassic of Poland" Acta Palaeontologica Polonica, doi:10.4202/app.2010.0045
Ortega, F., Escaso, F & Sanz, J.L. (2010) "A bizarre, humped Carcharodontosauria (Theropoda) from the Lower Cretaceous of Spain" Nature, 467, 203-206.
Ősi, A., Apesteguía, S. & Kowalewski, M. (2010) "Non-avian theropod dinosaurs from the early Late Cretaceous of Central Europe" Cretaceous Research, 31, 304-320.
Rauhut, O.W.M. (2003) "The interrelationships and evolution of basal theropod dinosaurs" Special Papers in Palaeontology, 69, 1-213.
Rauhut, O.W.M. (2007) "A fragmentary theropod skull from the Middle Jurassic of Patagonia" Ameghiniana, 44, 479-483.
Rauhut, O.W.M. (2011) "Theropod Dinosaurs from the Late Jurassic of Tendaguru (Tanzania)" Special Papers in Palaeontology, 86, 195-239. doi: 10.1111/j.1475-4983.2011.01084.x
Rauhut, O.W.M., Milner, A.C. & Moore-Fay, S. (2010) "Cranial osteology and phylogenetic position of the theropod dinosaur Proceratosaurus bradleyi (Woodward, 1910) from the Middle Jurassic of England" Zoological Journal of the Linnean Society, 158, 155-195.
Sadleir, R., Barrett, P.M. & Powell, H.P. (2008) "The anatomy and systematics of Eustreptospondylus oxoniensis, a theropod dinosaur from the Middle Jurassic of Oxfordshire, England" Monograph of the Palaeontographical Society, 627, 160, 1-82.
Smith, N.D., Makovicky, P.J., Pol, D., Hammer, W.R. & Currie, P.J. (2007) "Osteology of Cryolophosaurus ellioti (Dinosauria: Theropoda) from the Early Jurassic of Antarctica and implications for early theropod evolution" Zoological Journal of the Linnean Society, 151, 377-421.
Smith, N.D., Makovicky, P.J., Agnolin, F.L., Ezcurra, M.D., Pais, D.F. & Salisbury, S.W. (2008) "A Megaraptor -like theropod (Dinosauria: Tetanurae) in Australia: support for faunal exchange across eastern and western Gondwana in the Mid-Cretaceous" Proceedings of the Royal Society B, 275, 2085-2093.
Sullivan, C., Hone, D. W. E., Xu, X. & Zhang, F. (2010) "The asymmetry of the carpal joint and the evolution of wing folding in maniraptoran theropod dinosaurs" Proceedings of the Royal Society B, 277, 2027-2033.
Xu, X., Clark, J.M., Forster, C.A., Norell, M.A., Erickson, G.M., Eberth, D.A., Jia, C. & Zhao, Q. (2006) "A basal tyrannosauroid dinosaur from the Late Jurassic of China" Nature, 439, 715-718.
Xu, X., Clark, J.M., Mo J., Choiniere, J., Forster, C.A., Erickson, G.M., Hone, D.W.E., Sullivan, C., Eberth, D.A., Nesbitt, S., Zhao, Q., Hernandez, R., Jia, C., Han, F. & Guo, Y. (2009) "A Jurassic ceratosaur from China helps clarify avian digital homologies" Nature, 459, 940-944.
Yates, A.M. (2005) "A new theropod dinosaur from the Early Jurassic of South Africa and its implications for the early evolution of theropods" Palaeontologia Africana, 41, 105-122.
Inne:
Allain, R. & Bailleulm, A. (2009) "First revision of the theropod from the Toarcian of Wazzant (High Atlas Mountains, Marocco): the oldest known tetanurae?" 1st International Congress on North African Vertebrate Palaeontology Abstracts, 16-17. [abstrakt]
Chure, D.J. (2000) "A New Species of Allosaurus From the Morrison Formation of Dinosaur National Monument (Utah-Colorado) and a Revision of the Theropod Family Allosauridae" Dysertacja doktorska; Columbia University.
Longrich, N. (2002) "Systematics of Sinosauropteryx" Journal of Vertebrate Paleontology, 22, 80A. [abstrakt]
http://dml.cmnh.org/2011Oct/msg00220.html