


Jesteś w: Artykuły o teropodach
Karnozaury (Carnosauria)
Autor: Maciej Ziegler, korekta: Dawid Mika, Tomasz Sokołowski
Ostatnia aktualizacja: 1.12.2011 r.
Nomenklatura:
- Carnosauria (Huene, 1920)
>Allosaurus fragilis ~ Passer domesticus (zmodyfikowano z Padian i Hutchinson, 1997)
Dla tego kladu często, choć nieprawidłowo, używa się nazwy Allosauroidea (sensu Sereno, 1998). - Allosauroidea (Marsh, 1878 vide Currie i Zhao, 1994)
<Allosaurus fragilis & Sinraptor dongi (Holtz, Molnar i Currie, 2004; zmodyfikowano z Padian i Hutchinson, 1997) - Sinraptoridae (Currie i Zhao, 1994)
>Sinraptor dongi ~ Allosaurus fragilis v Monolophosaurus jiangi v Cryolophosaurus ellioti v Carcharodontosaurus saharicus (zmodyfikowano z Sereno, 1998)
Według wcześniejszej definicji Padiana i Hutchinsona (1997) (>Sinraptor dongi ~ Allosaurus fragilis) do Sinraptoridae mogą należeć też allozauroidy zaliczane do Carcharodontosauria. - Allosauridae (Marsh, 1878)
>Allosaurus fragilis ~ Sinraptor dongi v Monolophosaurus jiangi v Cryolophosaurus ellioti v Carcharodontosaurus saharicus (zmodyfikowano z Sereno, 1998)
Według wcześniejszej definicji Padiana i Hutchinsona (1997) (>Allosaurus fragilis ~ Sinraptor dongi) do Allosauridae mogą należeć też allozauroidy zaliczane do Carcharodontosauria. - Carcharodontosauria (Benson, Carrano i Brusatte, 2010)
>Carcharodontosaurus saharicus & Neovenator salerii ~ Allosaurus fragilis v Sinraptor dongi (Benson, Carrano i Brusatte, 2010)
Jest to synonim Carcharodontosauridae wg definicji używanych przed 2010 r. - Carcharodontosauridae (Stromer, 1931)
>Carcharodontosaurus saharicus ~ Neovenator salerii v Allosaurus fragilis v Sinraptor dongi (Benson, Carrano i Brusatte, 2010) - Carcharodontosaurinae (Stromer, 1931 vide Brusatte i Sereno, 2008)
<Carcharodontosaurus saharicus & Giganotosaurus carolinii (Brusatte i Sereno, 2008) - Giganotosaurinae (Coria i Currie, 2006)
>Giganotosaurus carolinii & Mapusaurus roseae ~ Carcharodontosaurus saharicus
Brusatte i Sereno (2008) zmienili nazwę na Giganotosaurini, aby czyniła zadość hierarchii linneuszowskiej oraz zdefiniowali ją inaczej - bardziej stabilnie: (<Giganotosaurus carolinii ~ Carcharodontosaurus saharicus). - Neovenatoridae (Benson, Carrano i Brusatte, 2010)
>Neovenator salerii ~ Carcharodontosaurus saharicus v Allosaurus fragilis v Sinraptor dongi (Benson, Carrano i Brusatte, 2010) - Megaraptora (Benson, Carrano i Brusatte, 2010)
>Megaraptor namunhuaiquii ~ Chilantaisaurus tashuikouensis v Neovenator salerii v Carcharodontosaurus saharicus v Allosaurus fragilis (Benson, Carrano i Brusatte, 2010) - Inne. Metriacanthosaurus prawdopodobnie jest przedstawicielem kladu Sinraptoridae, więc utworzona dla niego podrodzina Metriacanthosaurinae (Paul, 1988) ma - jako rodzina - pierwszeństwo przed Sinraptoridae, jest to jednak nazwa bez definicji filogenetycznej i praktycznie nieużywana.
Charakterystyka:
"Przybyły stworzenia jeszcze głodniejsze i trzykrotnie większe - karcharodontozaury. Dla nich wszystko, co żywe nadaje się do jedzenia"
Planeta dinozaurów: Jajo Alfy" (Dinosaur Planet: Alpha's Egg)
Karnozaury to grupa średniej do olbrzymiej wielkości aweteropodów, bardziej "konserwatywna" niż celurozaury. Ogólną budową przypominały bardziej bazalne tetanury. Mogły być częściowo opierzone (zob. opis Concavenator).
Ewolucja i budowa
Wśród Carnosauria wyróżnia się Allosauroidea, do którego należą wszystkie niewątpliwe karnozaury. Charakteryzują je m. in.: tylne zgrubienie (guz) na kości zaoczodołowej, obecność podoczodołowego wyrostka kości łzowej, zwrócone do przodu wyrostki kolczyste tylnych kręgów grzbietowych, szewrony ze środkowej części ogona w kształcie litery L, dystalne rozszerzenie kości łonowej w równym stopniu do przodu, jak i do tyłu oraz wyraźny tylnoprzyśrodkowy wyrostek na kości skokowej.
Allozauroidy miały ten sam, typowy plan budowy: duża, trójkątna i nieco wydłużona czaszka, często z ornamentacją w postaci rogów nad oczami, mocne, trójpalczaste kończyny przednie oraz długi ogon.
Karnozaury przeważnie trzymały głowę pochyloną - u akrokantozaura około 25 stopni w stosunku do poziomu. Kończyny tylne były mocno zbudowane. U większości z nich uda były nieco krótsze od podudzia, a śródstopie dwukrotnie krótsze od podudzia, co dowodzi tego, że nie były sprinterami (wyjątkiem zapewne były neowenatorydy). Nie miały też arctometatarsus, co czyniło je znacznie gorszymi biegaczami, niż uważany błędnie za powolnego Tyrannosaurus. Allozauroidy w niewielkim stopniu widziały obuocznie, podobnie jak dzisiejsze krokodyle. Budowa ich czaszek uniemożliwiała szerokie pokrywanie się pól widzenia prawego i lewego oka, co także jest swoistą ułomnością w porównaniu do tyranozaurydów. Nie miały także tak silnych szczęk, jak tyranozaur. Tak więc to raczej one powinny być uważane za padlinożerców, a jest całkiem inaczej. Uważa się je bowiem za przystosowane do zabijania, w tym także olbrzymich zauropodów (zob. "Dieta").
Karnozaury dzielą się na cztery grupy - bardziej bazalne, wcześniejsze (głównie albo raczej wyłącznie jurajskie) i uboższe w gatunki allozaurydy i sinraptorydy oraz przeważnie późniejsze (głównie kredowe), bardziej zaawansowane i liczniejsze karcharodontozaury, które z kolei dzieli się na karcharodontozaurydy i neowenatorydy. Prawdopodobnie od wspólnego pnia pierwsze oddzieliły się sinraptorydy, a następnie allozaurydy, choć pewne cechy łączą sinraptorydy i zaawansowane karcharodontozaury, np. budowa mózgoczaszki, obecność przedniej odnogi na kości jarzmowej i brak pneumatycznych otworów powiązanych z wewnętrzną tętnicą szyjną.
1. Sinraptoridae. Są one znane ze środkowej-późnej jury Chin i prawdopodobnie też Europy. Piszczel z Tajlandii może pochodzić z późnej jury lub wczesnej kredy (Buffetaut i Suteethorn, 2007). Azjatyckie, znane z materiału czaszkowego, sinraptorydy miały nozdrza zewnętrzne z wyraźną wstawką na tylnej krawędzi, znacznie większa okno promaxillary niż okno szczękowe i kołnierz kości łuskowej zakrywający głowę kości kwadratowej w widoku bocznym, ale nie miały rostralnej gałęzi kości szczękowej (za Holtz i in., 2004). Azjatyckich i europejskich przedstawiciele tej grupy łączą cechy obecne też u innych bazalnych tetanurów: stosunkiem długości do szerokości szypuły łonowej kości biodrowej wynoszącym 1,55-1,75 (obecne też u spinozauroidów), wąskim dołem na wewnętrznym wyrostku kości biodrowej z prawie równoległymi krawędziami (obecne też u niektórych megalozaurydów), bocznym kłykciem kości piszczelowej zbiegającym się z grzebieniem goleniowym w widoku z tyłu (obecne też u wielu megalozauroidów i Dilophosaurus) (za Benson, 2010). Miały też pewne cechy prymitywne, takie jak obecność IV kości śródręcza czy słabo rozwinięta dystalna ekspansja kości łonowej.
Rauhut (2003), na podstawie opublikowanych deskrypcji, zasugerował, że znane w tym czasie sinraptorydy (Sinraptor dongi, S. hepingensis, Yangchuanosaurus magnus, Y. shangyouensis) są tym samym gatunkiem, wskazując, że różnice między S. dongi a S. hepingensis są subtelne i mogą wynikać ze zmienności osobniczej; Holtz i in. (2004) wysnuli przypuszczenie, że Y. magnus i Y. shangyouensis to odowiednio starszy i młodszy osobnik tego samego gatunku.
2. Allosauridae. Podobnie jak sinraptorydy, nie są liczną grupą. Jedynym pewnym przedstawicielem jest dobrze zbadany, późnojurajski (kimeryd-tyton, 156-145 Ma) Allosaurus z Ameryki Północnej i Europy. Fragmentaryczne szczątki przypisywane allozaurydom znane są z późnej jury i wczesnej kredy Europy, Afryki i Azji.
3. Carcharodontosauria to grupa, której przedstawiciele mieli wiele charakterystycznych cech (gwiazdką oznaczono te, które ma Concavenator, reszta jest niepewna): czaszki (kości nosowe niemal jednakowej szerokości przez całą długość, jeden otwór Meckela w żuchwie), kręgów (silnia i charakterystyczna pneumatyzacja, wewnętrzna powierzchnia intercentrum obrotnika pochylona przednio-grzbietowo, grzbietowe hyposphenes formujące poprzecznie wąskie płaty) oraz kończyn tylnych (przedniotylna długość dystalnego rozszerzenia łonowego wynosząca ponad 60% długości trzonu łonowego, *mocno wklęsła biodrowa powierzchnia stawowa kości kulszowej, *pochylona tylno-przyśrodkowo głowa kości udowej, *wystająca niemal przyśrodkowo kostka przyśrodkowa dystalnej części kości piszczelowej, stosunek szerokości przednio-tylnej dystalnej kości strzałkowej do minimalnej szerokości trzonu mniejszy niż 1.7) (za Benson i in., 2010).
Dzieliły się na dwie grupy:
- Carcharodontosauridae. Znane są one z późnej jury (Veterupristisaurus) i "środkowej" kredy. Cechują się dodatkowymi otworami w kości szczękowej (także u sinraptorydów), zmniejszeniem okna przedoczodołowego, stykającymi się kośćmi zaoczodołowymi i łzowymi (obecne u Concavenator), wyrostkiem zaoczodołowym w formie małej ostrogi (u Acrocanthosaurus, Concavenator i bardziej zaawansowanych z nich jest to już duży, zakrzywiony kołnierz) i przedniowewnętrznym końcem rowka przyzębowego nachylonym przedniowewnętrznie. Cechy nieznane u Concavenator i Eocarcharia to: rozrośnięty dorsowentralnie i rozdzielony anterowentralnym ('podbródkowym') wyrostkiem przedni koniec kości zębowej, wyraźny wzdłużny rowek około środka wysokości kości zębowej - na jej bocznej powierzchni oraz wysokie i poprzecznie szerokie łuki kręgowe na kręgach grzbietowych z centralnymi regionami powierzchni bocznej otoczonymi głębokimi rowkami (tworzące strukturę w kształcie dwuteownika) (za Benson i in., 2010). Kończyny przednie były dość krótkie. Duże guzy nad oczami, obecne też u abelizaurydów i niektórych tyranozaurydów, mogły służyć do uderzania się głowami.
Wśród karcharodontozaurydów wyróżnia się ubogie w gatunki oraz ograniczone czasowo i geograficznie (ale za to zawierające największe karnozaury), wyspecjalizowane klady:
- Carcharodontosaurinae to grupa olbrzymich karcharodontozaurydów z cenomanu (100-93 Ma) Gondwany (Afryka i Ameryka Płd.). Synapomorfie karcharodontozaurynów (być może występujące też, prócz ostatniej, u Tyrannotitan) to: mocno rzeźbione kości szczękowe (obecne też u Neovenator, którego czaszka ogólnie jest mniej pomarszczona), przedniośrodkowy róg górnego dołu skroniowego kości czołowej przykryty półką czołowociemieniową, kąt między osią kłykcia potylicznego i poprzeczną płaszczyzną basal tubera mniejszy niż 75 stopni, skostnienie przegrody międzyoczodołowej oraz prosty profil tylnej krawędzi zębów mieszczących się pośrodku linii zębów (za Brusatte i Sereno, 2008; Sereno i Brusatte, 2008; Brusatte i in., 2009). Należą do nich rodzaj Carcharodontosaurus i klad:
-> Giganotosaurinae to południowoamerykańska linia zaawansowanych, kolosalnych karcharodontozaurynów - Giganotosaurus i prawdopodobnie także Mapusaurus. Łączy je zaawansowana budowa kości udowej: niski czwarty krętarz (obecne też, lecz bardziej wyraźnie, u Chilantaisaurus i wielu celurozaurów) i szeroki rowek prostownika na dystalnym końcu tej kości (taką budowę ma też Tyrannotitan, może on więc być wczesnym giganotozaurynem). Giganotosaurus i Mapusaurus miały silniej niż Carcharodontosaurus (i zapewne też Concavenator) pomarszczone kości nosowe - były one pokryte charakterystycznymi naroślami. - Neovenatoridae to późno rozpoznana (opublikowana przed drukiem w 2009 r.) i zróżnicowana grupa nietypowych karcharodontozaurów mająca cechy celurozaurów (np. pneumatyzacja obręczy - miednicznej i barkowej, co jest obecne też u Piatnitzkysaurus). Pozostają dość słabo znane, gdyż większość ich przedstawicieli jest dość fragmentaryczna. Odróżnia je od innych teropodów kilka cech kręgów (tylne ogonowe z małymi, kołnierzastymi bocznymi rozszerzaniami na stronie zazygapofyzjalnej), kończyn (stosunek długości do najmniejszej szerokości łopatki między 7, 5 a 9, biodrowy przedstawowy dół ograniczony grzbietowomedialnie przez wyraźną półkę, przednioboczny wyrostek bocznego kłykcia piszczeli zakręca przyśrodkowo jako szpiczasty wyrostek, boczny kłykieć kości udowej skierowany bardziej ku tyłowi niż środkowy - co jest obecne także u Carcharodontosaurus, silnie spneumatyzowana kość biodrowa) oraz być może też czaszki (np. inaczej niż u innych karnozaurów przedni wyrostek kości zaoczodołowej gładki i nierozrośnięty poprzecznie, co zaobserwowano u Aerosteon i Orkoraptor), która jest w większości znana tylko u Neovenator - najbardziej bazalnego z nich - a u bardziej zaawansowanych znane są jedynie fragmenty. Neovenator ogólną budową był podobny do bardziej bazalnych tetanurów np. Allosaurus i zapewne zajmował podobną niszę ekologiczną, czego nie da się powiedzieć o przedstawicielach kladu:
- Megaraptora to grupa zaawansowanych neowenatorydów o unikalnej budowie, niespotykanej u innych niecelurozarowych tetanurów. Miały wydłużone, potężne przednie łapy a na nich wielkie i wąskie pazury - zapewne przystosowane do zabijania (być może to także cecha neowenatora). Umiały szybko biegać, na co wskazują wydłużone dolne partie kończyn - kości śródstopia były długie i smukłe. Miały one wysoki mniejszy krętarz udowy i zmodyfikowany staw skokowy z długi, wyrostkiem wstępującym kości skokowej i wyraźną przyporą skokową na piszczeli. Cechuje je też znaczny stopień pneumatyzacji kości kończyn. Nieopublikowana analiza Mortimera (online) wskazuje, że są one celurozaurami.
Generalnie karnozaury były podobne do bardziej bazalnych tetanurów. Wiele teropodów uznawanych często za bardziej bazalne tetanury może należeć do omawianej grupy (Afrovenator, Becklespinax, Erectopus, Lourinhanosaurus, Marshosaurus, Metriacanthosaurus, Monolophosaurus, Piatnitzkysaurus, Poekilopleuron, Shidaisaurus, Sigilmassasaurus); dotyczy to w mniejszym stopniu także celurozaurów (Gasosaurus, Iliosuchus). Analiza Rauhuta (2003) wskazuje, że teropody zaliczane powszechnie do Carnosauria albo Megalosauroidea tworzą jeden, obszerniejszy klad niecelurozaurowych tetanurów. Niecodzienną topologię przedstawia też wynik drugiej analizy Xu i współpracowników (2009). Karnozaurami są tam tylko allozaurydy i sinraptorydy, natomiast obszerny klad bardziej bazalnych tetanurów (odpowiednia nazwa to Carcharodontosauria) tworzą: karcharodontozauryny (w tym Acrocanthosaurus), które są grupą siostrzaną kladu Eustreptospondylus+Dubreuillosaurus (uważanych raczej za megalozaurydy), spinozauroidy (Torvosaurus+Spinosauridae) oraz Neovenator (jako najbardziej bazalny przedstawiciel Carcharodontosauria). Jedna z analiz (Brusatte i in., 2009 - I) wskazuje, że poszczególne grupy wliczane do Carnosauria nie tworzą niewątpliwie kladu, lecz politomię wraz z Coelurosauria, ale jak później zaznaczają ci sami autorzy (Brusatte i in., 2010A), w analizie ilość cech ważnych dla tej części drzewa jest ograniczona. Podobny wynik mają analizy Choiniere i in. (2010A i 2010B), gdzie Sinraptor jest celurozaurem. Jednak większość badań wskazuje, że karnozaury są odrębną, bardziej zaawansowaną (bliższą ptakom) od Megalosauroidea i monofiletyczną grupą.
Dieta
Wiele cech budowy karnozaurów interpretuje się jako przystosowania do polowania na duże zauropody, które były często dominującymi roślinożercami w ekosystemach zasiedlonych przez omawiane teropody. Znane są ślady zębów oraz zgubione zęby allozaura w kościach zauropodów, stegozaura a także torwozaura. W Teksasie znaleziono odciski stóp pozostawione przez Acrocanthosaurus (lub pokrewną formę), który prawdopodobnie zaatakował dużego zauropoda. Karnozaury, mając do dyspozycji mniejszą i łatwiejszą do upolowania zdobycz, zapewne nie ograniczały się tylko do zauropodów, jednak należy zauważyć, że wśród większości z nich nie zaobserwowano przystosowań do szybkiego biegu, widocznego wśród innych grup teropodów (ceratozaurów, celurozaurów) - a jak wiadomo - zauropody nie były mistrzami prędkości.
Kończyny przednie prawdopodobnie były pomocne w przytrzymywaniu ofiary (więcej o roli kończyn w opisie Acrocanthosaurus), jednak główną bronią większości karnozaurów, a szczególnie zaawansowanych karcharodontozaurydów, była czaszka. Analiza budowy allozaura, wykazująca stosunkowo słabą siła ścisku szczęk, które mogły być otwieranie niezwykle szeroko i mocno zbudowaną czaszka skłoniły badaczy do przypuszczenia, że ten karnozaur polował na zauropody, rzucając się na nie z szeroko rozdziawionymi szczekami, które używał niczym topora i ciął nimi swoje wielkie ofiary (Bakker, 2000, Rayfield i in., 2001). Taki sposób polowania jest raczej mało prawdopodobny, ponieważ żuchwa mogła ulec zwichnięciu, a szyja byłaby zbyt mocno wygięta do góry. Prawdopodobnie allozaury gryzły zauropody w wypukłe części ciała, a potężne mięśnie szyi kierowały głowę w bok, a nie do góry i w dół (Antón i in., 2003). Taki sposób polowania był bardziej prawdopodobny, ponieważ zaatakowane ofiary na pewno próbowały uciec, a przynajmniej zmienić pozycję. Zapewne był też bezpieczniejszy dla samych allozaurów.
Wyjątkiem były neowenatorydy - szczególnie megaraptory, których budowa sugeruje szybkobieżność i zabijanie ofiar za pomocą olbrzymich pazurów kończyn przednich.
W brzusznej okolicy Poekilopleuron - prawdopodobnie wczesnego sinraptoryda - znaleziono szczątki ryb chrzęstnoszkieletowych, w tym ząb rekina Polyacrodus. Co więcej, zachowanie się delikatnych kości jego szkieletu (np. gastralia), wydobytego z morskich osadów, wskazuje na to, że po śmierci ciało poekilopleurona nie było transportowane z lądu.
Wymiary:
?~ 4 m (Fukuiraptor), 5-6 m (?Australovenator, Shaochilong) <<< 13-13,5 m (Giganotosaurus)
Teropody, które mogą być bazalnymi karnozaurami lub ich bliskimi krewnymi (Monolophosaurus, Gasosaurus) mierzą średnio 4-5 m i ważą 200-500 kg. Bardziej zaawansowane (i niewątpliwe) jurajskie karnozaury były przeważnie już sporo większe (6-9 m, 0,5-2 t - prócz mierzącego 4-5 m Lourinhanosaurus, którego w pełni wyrośnięty osobnik osiągał zapewne sporo więcej). Duży był jurajski karcharodontozauryd Veterupristisaurus (8,5-10 m), ale to niektóre późnojurajskie allozaurydy (Saurophaganax, 10-12 m, 3 t) i sinraptorydy (Yangchuanosaurus magnus, 10-11 m, 3 t) osiągały już olbrzymie rozmiary. Z późnego oksfordu (161-158 Ma) Chin pochodzi największy ząb jurajskiego teropoda, prawdopodobnie sinraptoryda - jego korona mierzy 9,2 cm (Xu i Clark, 2008). Porównując rozmiary zęba do zębów Y. magnus, długość ciała tajemniczego teropoda można obliczyć na ponad 14 m! Pomijając właściciela owego zęba, to liczne kredowe karcharodontozaury były prawdziwymi gigantami - Chilantaisaurus, Acrocanthosaurus i Tyrannotitan osiągały 11-12 m długości i 2,5-6 t masy (ten ostatni ma wg Paula (2010) aż 13 m i 7 t). Jeszcze większe były karcharodontozauryny - wszystkie z nich osiągały ponad 12 m i 4-10 t (Carcharodontosaurus iguidensis mógł być mniejszy). Obok nich żyły formy dużo mniejsze, prawdopodobnie wtórnie zminiaturyzowane - klad Fukuiraptor + Australovenator oraz karcharodontozauryd Shaochilong. Masa neowenatorydów wahała się od ok. 175 kg (Fukuiraptor) do 2,5-6 t (Chilantaisaurus). Większość karnozaurów (prócz największych karcharodontozaurydów) zapewne mieściła się w tym przedziale.
Kiedy:
~ 166 Ma -> 72-68 Ma
najstarsze:
? Poekilopleuron 167-165 Ma
? Shidaisaurus 176-168 Ma
Sinraptor 161-158 Ma
najmłodsze: Orkoraptor 72-68 Ma
Najstarszy pewny karnozaur znany jest dopiero z późnej jury (Sinraptor dongi). Kilka form, które mogą być karnozaurami, pochodzi z wcześniejszego przedziału czasowego (Gasosaurus i Monolophosaurus - ?165-161 Ma, Poekilopleuron - 167-166 Ma, Shidaisaurus - 176-168 Ma). W każdym razie karnozaury oddzieliły się od pozostałych aweteropodów (celurozaurów) najpóźniej w środkowej jurze, a na początku późnej były już szeroko rozpowszechnione.
Karcharodontozaury na pewno pojawiły się w pierwszej połowie późnej jury. Najwcześniejszym z nich jest afrykański karcharodontozauryd Veterupristisaurus z formacji Tendaguru (późny kimeryd, 153-150 Ma) a kolejne szczątki należące być może do tej grupy to kilka niekompletnych zębów z tytonu (151-145 Ma) Etiopii wykazujących podobieństwo do Acrocanthosaurus (Goodwin i in., 1999) oraz zęby z późnej jury (?oksford-późny tyton: ?161-145 Ma) Tanzanii (Megalosaurus ingens) (Rauhut, 1995; 2011). Saurophaganax, powszechnie uważany za allozauryda, według analizy Cau (online) jest bazalnym karcharodontozaurem. Concavenator - prawdopodobnie karcharodontozauryd pochodzi z późnego barremu (128-125 Ma). W każdym razie najwcześniejszym dość kompletnym karcharodontozaurydem jest Tyrannotitan z aptu (125-112 Ma) Argentyny, a neowenatorydem - bazalny Neovenator z barremu (130-125 Ma) Anglii lub bardziej zaawansowany Fukuiraptor z wczesnej kredy - prawdopodobnie także barremu. Karcharodontozaurydy we wczesnej kredzie były pospolite i dotrwały przynajmniej do cenomanu (~94 Ma), a być może nawet mastrychtu (71-65 Ma) natomiast neowenatorydy na pewno do wczesnego mastrychtu (72-68 Ma). Do wczesnej kredy dotrwały być może allozaurydy. Karcharodontozaury były dominującymi drapieżnikami na wielu kontynentach do późnej kredy (?cenoman, ?turon). Na końcu kredy były rzadsze, lecz neowenatorydy stanowiły wciąż ważny składnik fauny.
Gdzie:
Laurazja (Ameryka Płn., Azja, Europa) i Gondwana (Afryka, Ameryka Płd., Australia)
Karnozaury pojawiły się, gdy dzisiejsze kontynenty tworzyły jeszcze jeden ląd, więc zapewne zamieszkiwały one całą planetę. W późnej jurze częstsze były na północy (allozaurydy - Ameryka Płn. i Europa, sinraptorydy - Azja i prawdopodobnie Europa) i prawdopodobnie stamtąd pochodziły. Bazując na wynikach analizy Bensona i współpracowników (2010) najbardziej prawdopodobnym miejscem genezy karnozaurów jest Europa (co ciekawe sinraptorydy są grupą europejską, z jednym azjatyckim zaawansowanym kladem), jednak może to wynikać z niekompletności zapisu kopalnego (zob. też artykuł o celurozaurach). Karcharodontozaury cechuje ogólnoświatowy zasięg, lecz najwcześniejsze i najpóźniejsze z nich ograniczały się do Gondwany. Karcharodontozaury one ważnymi, o ile nie dominującymi, drapieżnikami na wielu kontynentach we wczesnej i na początku późnej kredy - być może aż do kampanu (Ameryka Płd. i Płn., Europa, Afryka, Azja - ?Siamotyrannus, Kelmayisaurus). Królestwem neowenatorydów prawdopodobnie była kredowa Australia, gdzie znaleziono kilku przedstawicieli tej grupy. Wczesne i bazalne karcharodontozaury znane są z Europy, więc być może stamtąd pochodzą karcharodontozaurydy i neowenatorydy.
Być może megaraptory zajmowały podobne nisze ekologiczne co celurozaury (należy zauważyć, że nie występowały w Ameryce Płn. i Azji, gdzie celurozaury dominowały), co pozwalało im zmniejszyć konkurencję ze strony karcharodontozaurydów (po których wymarciu w Ameryce Płd. osiągnęły duże rozmiary).
Dowiedz się więcej:
- o karnozaurach o niepewnej klasyfikacji:
??Becklespinax, ??Erectopus, ??Shidaisaurus, Siamotyrannus, ??Valdoraptor
- sinraptorydach:
(?)Leshansaurus, ??Lourinhanosaurus, ?Metriacanthosaurus, ??Poekilopleuron, Sinraptor, (Szechuanosaurus), Yangchuanosaurus
- allozaurydach:
Allosaurus, "Madsenius", (?)Saurophaganax, "Wyomingraptor"
- bazalnych karcharodontozaurydach:
?Acrocanthosaurus, ?Concavenator, ?Eocarcharia, ?Kelmayisaurus, ?Shaochilong, Veterupristisaurus
- karcharodontozaurynach:
Carcharodontosaurus, Giganotosaurus, Mapusaurus, ??Tyrannotitan
- bazalnych neowenatorydach:
Chilantaisaurus, Neovenator
- (?) i megaraptorach:
Aerosteon, Australovenator/Rapator, Fukuiraptor, Megaraptor, Orkoraptor

Czaszka Acrocanthosaurus w widakach: od góry i z boku (z Currie i Carpenter, 2000)

Rekonstrukcja szkieletu Allosaurus (autor: Scott Hartman; rysunek pochodzi z tej strony).

Zakres otwierania paszczy u Allosaurus (autor: Stephen O'Connor; rysunek pochodzi z tej strony).
{kind=link}

Rekonstrukcja Neovenator salerii. Autor: Alexander Lovegrove. Rysunek pochodzi z tej strony.

Kladogram z Coria i Currie, 2006 (na podstawie Currie i Carpenter, 2000 z dodanym Mapusaurus i kilkoma drobnymi zmianami).

Kladogram z Brusatte i in., 2009 - I (niekompletny; na podstawie Smith i in., 2007 z dodanym Shaochilong).

Skalibrowany stratygraficznie kladogram z Hocknul i in., 2009 (na podstawie Brusatte i Sereno, 2008 z dodanym Australovenator).

Kladogram z Sereno i Brusatte, 2008 (na podstawie Brusatte i Sereno, 2008 z usunięciem 26 nieinformatywnych dla zbadania relacji między karcharodontozaurydami cech i taksonów, usunięciem 19 kolejnych i dodaniem 6 nowych).
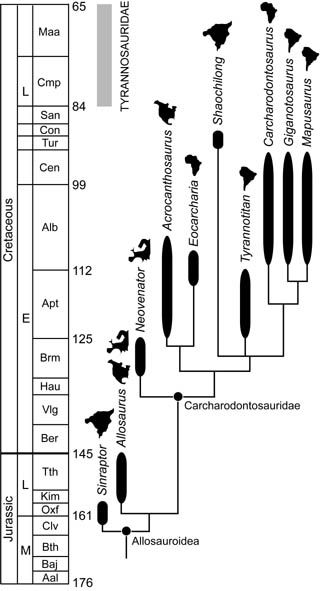
Skalibrowany stratygraficznie kladogram z Brusatte i in., 2009 - II (na podstawie Brusatte i Sereno, 2008 z dodanym Shaochilong, niektórymi poprawkami z Sereno i Brusatte, 2008, usunięciem 19 cech i dodaniem 6 nowych), ukazujący też miejsce znalezienia rodzajów i okres dominacji tyranozaurydów w Azji i Ameryce Płn. (szary prostokąt).

Kladogram z Eddy i Clarke, 2011.
Źródła:
Publikacje naukowe:
Agnolin, F.L., Ezcurra, M.D., Pais, D.F. & Salisbury, S.W. (2010) "A reappraisal of the Cretaceous non-avian dinosaur faunas from Australia and New Zealand: evidence for their Gondwanan affinities" Journal of Systematic Palaeontology, 8, 257-300
Allain, R. (2005A) "The postcranial anatomy of the megalosaur Dubreuillosaurus valesdunensis (Dinosauria Theropoda) from the Middle Jurassic of Normandy, France" Journal of Vertebrate Paleontology, 25, 850–858.
Allain, R. (2005B) "The enigmatic theropod dinosaur Erectopus superbus (Sauvage, 1882) from the Lower Albian of Louppy-le-Château (Meuse, France)" [w:] Carpenter, K. "The Carnivorous Dinosaurs" Indiana University Press, 72-86.
Antón, M., Sanchez, I.M., Salesa, M.J. & Turner, A. (2003) "The Muscle-Powered Bite of Allosaurus (Dinosauria; Theropoda): an Interpretation of Cranio-Dental Morphology" Estudios Geologicos, 59, 313-323.
Benson, R.B.J. (2010) "The osteology of Magnosaurus nethercombensis (Dinosauria, Theropoda) from the Bajocian (Middle Jurassic) of the United Kingdom and a re-examination of the oldest records of tetanurans" Journal of Systematic Palaeontology, 8, 131-146.
Benson, R.B.J., Carrano, M.T. & Brusatte, S.L. (2010) "A new clade of archaic large-bodied predatory dinosaurs (Theropoda: Allosauroidea) that survived to the latest Mesozoic" Naturwissenschaften, 97, 71-78.
Benson, R.B.J. & Barrett, P.M. (2009) "Dinosaurs of Dorset: Part I, the carnivorous dinosaurs (Saurischia, Theropoda)" Proceedings of the Dorset Natural History and Archaeological Society, 130, 133-147.
Benson, R.B.J. & Xu, X. (2008) "The anatomy and systematic position of the theropod dinosaur Chilantaisaurus tashuikouensis Hu, 1964 from the Early Cretaceous of Alanshan, People's Republic of China" Geological Magazine, 145,778-789.
Brusatte, S.L. & Sereno, P.C. (2008) "Phylogeny of Allosauroidea" Journal of Systematic Paleontology 6, 155-182.
Brusatte, S.L., Benson, R.B.J & Hutt, S. (2008) "The osteology of Neovenator salerii (Dinosauria: Theropoda) from the Wealden Group (Barremian) of the Isle of Wight" Monograph of the Palaeontographical Society, 162, 1-166.
Brusatte, S., Benson, R., Chure, D., Xu, X., Sullivan, C., & Hone, D. (2009) "The first definitive carcharodontosaurid (Dinosauria: Theropoda) from Asia and the delayed ascent of tyrannosaurids" Naturwissenschaften, 96, 1051-1058.
Brusatte, S.L., Chure, D.J., Benson, R.B.J. & Xu, X. (2010A) "The osteology of Shaochilong maortuensis, a carcharodontosaurid (Dinosauria: Theropoda) from the Late Cretaceous of Asia" Zootaxa 2334, 1-46.
Brusatte, S. L., Benson, R.B.J. & Xu, X. (2010B) "The evolution of large-bodied theropod dinosaurs during the Mesozoic in Asia" Journal of Iberian Geology, 36(2), 275-296.
Brusatte, S.L., Benson, R.B.J. & Xu, X. (2011) "A reassessment of Kelmayisaurus petrolicus, a large theropod dinosaur from the Early Cretaceous of China" Acta Palaeontologica Polonica, doi:10.4202/app.2010.0125
Cau, A., Dalla Vecchia, F.M. & Fabbri, M. (2011) "Evidence of a new carcharodontosaurid from the Upper Cretaceous of Morocco" Acta Palaeontologica Polonica, doi:10.4202/app.2011.0043
Choiniere, J.N., Xu, X., Clark, J.M., Forster, C.A., Guo, Y. & Han, F. (2010A) "A Basal Alvarezsauroid Theropod from the Early Late Jurassic of Xinjiang, China" Science, 327, 571-57
Choiniere, J.N., Clark, J.M., Forster, C.A. & Xu, X. (2010B) "A basal coelurosaur (Dinosauria: Theropoda) from the Late Jurassic (Oxfordian) of the Shishugou Formation in Wucaiwan, People's Republic of China" Journal of Vertebrate Paleontology, 30(6), 1773-1796. doi: 10.1080/02724634.2010.520779
Coria, R. A. & Currie P. J. (2006) "A new carcharodontosaurid (Dinosauria, Theropoda) from the Upper Cretaceous of Argentina" Geodiversitas, 28, 71-118.
Eddy, D.R. & Clarke, J.A. (2011) "New Information on the Cranial Anatomy of Acrocanthosaurus atokensis and Its Implications for the Phylogeny of Allosauroidea (Dinosauria: Theropoda)" PLoS ONE 6(3), e17932. doi:10.1371/journal.pone.0017932
Goodwin, M.B., Clemens, W.A., Hutchison, J.H., Wood, C.B., Zavada, M.S., Kemp, A., Duffin, C.J. & Schaff, C.R. (1999) "Mesozoic Continental Vertebrates with Associated Palynostratigraphic Dates from the Northwestern Ethiopian Plateau" Journal of Vertebrate Paleontology, 19, 728-741.
Han, F., Clark, J.M., Xu, X., Sullivan, C., Choiniere, J. & Hone, D.W.E. (2011) "Theropod teeth from the Middle-Upper Jurassic Shishugou Formation of northwest Xinjiang, China" Journal of Vertebrate Paleontology, 31(1), 111-126. DOI: 10.1080/02724634.2011.546291
Hocknull, S.A., White, M.A., Tischler, T.R., Cook, A.G., Calleja, N.D., Sloan, T. & Elliott, D.A. (2009) "New mid-Cretaceous (latest Albian) dinosaurs from Winton, Queensland, Australia" PLoS ONE, 4 e6190.
Holtz, T.R., Molnar, R.E. & Currie, P.J. (2004) "Basal Tetanurae" [w:] Weishampel, D.B., Dodson, P. & Osmólska, H. "The Dinosauria" Berkeley and Los Angeles: University of California Press, 71-110.
Naish, D. & Martill, D. M. (2007) "Dinosaurs of Great Britain and the role of the Geological Society of London in their discovery: basal Dinosauria and Saurischia" Journal of the Geological Society, London, 164, 493-510.
O'Connor, P. M. (2007) "The postcranial axial skeleton of Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar" [w:] Sampson, S. D. & Krause, D. W. (ed.) "Majungasaurus crenatissimus (Theropoda: Abelisauridae) from the Late Cretaceous of Madagascar" Society of Vertebrate Paleontology Memoir 8, 127-162.
Ortega, F., Escaso, F & Sanz, J.L. (2010) "A bizarre, humped Carcharodontosauria (Theropoda) from the Lower Cretaceous of Spain" Nature, 467, 203-206.
Rauhut, O. W. M. (1995) "Zur systematischen Stellung der afrikanischen Theropoden Carcharodontosaurus Stromer 1931 und Bahariasaurus Stromer 1934" Berliner geowissenschaftliche Abhandlungen. Reihe E: Palaeobiologie, 16, 357-375 [tłumaczenie na angielski: N. D. Smith; paleoglot.org]
Rauhut, O.W.M. (2003) "The interrelationships and evolution of basal theropod dinosaurs" Special Papers in Palaeontology, 69, 1-213.
Rauhut, O.W.M. (2011) "Theropod Dinosaurs from the Upper Jurassic of Tendaguru (Tanzania)" Special Papers in Palaeontology, 86, 195-239.
Rauhut, O. W. M. & Lopez-Abarello, A.(2008) "Archosaur evolution during the Jurassic: a southern perspective" Revista de la Asociación Geológica Argentina, 63, 557-585.
Sereno, P.C. & Brusatte, S.L. (2008) "Basal abelisaurid and carcharodontosaurid theropods from the Lower Cretaceous Elrhaz Formation of Niger" Acta Palaeontologica Polonica, 53, 15-46.
Xu, X. & Clark, J.M. (2008) "The presence of a gigantic theropod in the Jurassic Shishugou Formation, Junggar Basin, western China" Vetebrata PalAsiatica, 46, 157-160.
Xu, X., Clark, J.M., Mo J., Choiniere, J., Forster, C.A., Erickson, G.M., Hone, D.W.E., Sullivan, C., Eberth, D.A., Nesbitt, S., Zhao, Q., Hernandez, R., Jia, C., Han, F. & Guo, Y. (2009) "A Jurassic ceratosaur from China helps clarify avian digital homologies" Nature, 459, 940-944.
Inne:
http://theropoddatabase.blogspot.com/20 ... ridae.html
Paul, G.S. (2010) "The Princeton Field Guide to Dinosaurs" wyd. Princeton University
http://www.forum.dinozaury.com/posting. ... =6&p=33956