


Jesteś w: Artykuły o teropodach
Teropody (Theropoda) bazalne
Autorzy: Maciej Ziegler, Dawid Mika
Ostatnia aktualizacja: 19.11.2011 r.
Nomenklatura:
- Theropoda (Marsh, 1881)
>Allosaurus fragilis ~ Morosaurus impar (zmodyfikowano z Kischlat, 2000) - Herrerasauria (Galton, 1985)
>Herrerasaurus ischigualastensis ~ Liliensternus liliensterni v Plateosaurus engelhardti (zmodyfikowano z Langer, 2004) - Herrerasauridae (Benedetto, 1973)
<Herrerasaurus ischigualastensis & Staurikosaurus pricei (zmodyfikowano z Novas, 1992)
Do tak definiowanego kladu mogą należeć niemal wszystkie teropody (Nesbitt i in., 2011). - Avepoda (Paul, 2002)
> I kość śródstopia niestykająca się z dystalnymi kośćmi stępu (Passer domesticus) (zmodyfikowano z Paul, 2002)
Jest to klad o zasięgu bardzo zbliżonym do Neotheropoda, lecz teoretycznie szerszym: Tawa wydaje się nie być członkiem tego kladu, ale celofyzoidy już tak. W matrycy Sereno, gdzie Tawa jest celofyzoidem (opublikowanej w Martinez i in., 2011), nie ma tej cechy. - Neotheropoda (Bakker, 1986)
<Coelophysis bauri & Passer domesticus (zmodyfikowano z Sereno, 1998)
Bakker pierwotnie użył tej nazwy dla określenia teropodów bardziej zaawansowanych od Podokesauridae (?=Coelophysidae, zob. artykuł o Coelophysoidea). - Averostra (Paul, 2002)
<Ceratosaurus nasicornis & Allosaurus fragilis (Ezcurra i Cuny, 2007)
Pierwotna definicja Paula jest bardzo problematyczna: > dodatkowe okno w kości szczękowej (promaxillary fenestra) (Dromaeosaurus albertensis). Homologia tego okna jest jednak trudna do stwierdzenia, gdyż obecność otworów jest prawdopodobnie wysoce homoplastyczna. Taki otwór obecny jest np. u Eodromaeus, Herrerasaurus, Sanjuansaurus, Pampadromaeus (dół), Zupaysaurus, Heterodontosaurus ale nie u Daemonosaurus, Eoraptor i Tawa. Wg Martineza i in. (2011) jest to synapomorfia Theropoda, utracona u celofyzoidów.
Charakterystyka:
Ewolucja: pierwsze teropody
Teropody należą do dinozaurów gadziomiednicznych (Saurischia), a ich najbliższymi krewnymi, jak już wynika z definicji filogenetycznej, są zauropodomorfy, z którymi tworzą grupę Eusaurischia. Prawdopodobnie przodkowie teropodów były mięsożernymi, dwunożnymi zwierzętami a hipoteza, że byli wszystkożerni i być może fakultatywnie czworonożni, jest minimalnie mniej prawdopodobna (Nesbitt i in., 2010; Barrett i in., 2011). Możliwe, że byli pokryci protopiórami.
Pozycja filogenetyczna domniemanych wczesnych teropodów - herrerazaurów i Eoraptor - pozostaje kwestią sporów - negowano nawet ich przynależność do dinozaurów, choć najnowsze i najobszerniejsze dotąd badanie (Nesbitt i in., 2009 i oparte na nim: Suesa i in., 2011, Ezcrurry i Brusattego, 2011 oraz Langera i in., 2011) wskazuje, że są to jednak najbardziej bazalne teropody. Podobne kontrowersje dotyczą Guaibasaurus, który, tak jak Eoraptor, może być zauropodomorfem.
Cechy łączące niewątpliwe teropody z herrerazaurami i eoraptorem często uważa się za wynikające z podobnego sposobu odżywiania (drapieżności), lecz należy zauważyć, że także wiele z nich nie wynika jedynie z podobnego trybu życia. Synapomorfie Theropoda to (głównie za Nesbitt i in., 2009; gwiazdką oznaczono obecne u eoraptora):
- wgłębienie w kości klinowej podstawowej*,
- staw panewkowy żuchwy ulokowany na poziomie dorsalnej krawędzi kości zębowej*,
- ostre, zakrzywione, spłaszczone bocznie i ząbkowane zęby,
- nierozrośnięte środkowodystalnie korony zębów,
- wydłużone prezygapofyzje na dystalnych kręgach ogonowych,
- skrócona kość ramieniowa (poniżej 60 % długości udowej)*,
- stykające się (lecz nie zachodzące na siebie) proksymalne końce kości śródręcza*,
- głębokie i asymetryczne wgłębienia prostownika na proksymalnodorsalnej części I-III kości śródręcza*,
- trzon IV kości śródręcza znacznie węższy niż u pozostałych*,
- wydłużone końcowe paliczki dłoni,
- środkowobocznie cienka dystalna krawędź kości łonowej,
- kłykieć boczny (strzałkowy) proksymalnej części kości piszczelowej na poziomie środkowego kłykcia na jego tylnym obrzeżu*
- staw śródżuchwowy, dzięki sporemu (~15 stopni w górę i w dół) zakresowi ruchu ułatwiający prawdopodobnie łapanie zdobyczy, obecny u eoraptora, neoteropodów i ?herrerazaurów. Staw mógł pomagać w połykaniu dużych ofiar lub amortyzować uderzenia.*
Pozycja filogenetyczna Herrerasauria wśród dinozaurów jest niepewna - prawdopodobnie są bazalnymi przedstawicielami Theropoda lub Saurischia. Niektóre podobieństwa do teropodów mogą wynikać z podobnego sposobu odżywiania - kwesta ta wymaga bardziej obszernych badań. Wśród nich wyróżnią się jeszcze Herrerasauridae. Herrerazaury występowały w obu Amerykach, w Indiach oraz w Europie, choć jedyne niewątpliwe szczątki pochodzą z Ameryki Południowej. Miały duże głowy i były niewątpliwe mięsożerne (w przeciwieństwie do eoraptora, który mógł być wszystkożerny). Koniec ogona był usztywniony, co działało stabilizująco podczas biegu (podobnie jak u tetanurów). Herrerazaury łączą następujące cechy: brak wyraźnego zagłębienia dla przyczepu mięśnia caudifemoralis (rewersja ze stanu stwierdzonego u kladu Silesauridae+Dinosauria, obecne też u zauropodomorfów bardziej zaawansowanych od Saturnalia), brak rowka na grzbietowobocznej powierzchni bliższej części kości kulszowej (rewersja ze stanu stwierdzonego u Dinosauria lub obecnego już wcześniej) i obecność półki przedniego kłykcia proksymalnej do czwartego kłykcia na kości udowej (miejsce przyczepu dla mięśnia iliofemoralis externus; obecne też u celofyzoidów) (za Nesbitt i in., 2009). Teropody zaliczane do Herrerasauria mogą tworzyć grad (Nesbitt i in., 2011).
Ewolucja: neoteropody
Ponad wszelką wątpliwość do Theropoda należą cztery taksony - rodzaje Daemonosaurus, Eodromaeus i Tawa oraz grupa Neotheropoda (Tawa może być neoteropodem). Wyróżnia się też klad Avepoda, do którego należą wszystkie neoteropody. Ich charakterystyczne cechy to (za Nesbitt i in., 2009):
- cienki tylnogórny wyrostek kości przedszczękowej,
- wentralny wyrostek na tylnym końcu głównej części kości przedszczękowej,
- szew kości przedszczękowej i nosowej w kształcie litery W,
- głębokość wewnętrznej części okna przodoczodołowego dużo większa niż głębokość kości szczękowej poniżej wewnętrznej krawędzi okna przodoczodołowego,
- wewnętrzna gałąź opisthotic (= crista interfenestralis) zakryta przez najbardziej boczną krawędź kości bocznopotylicznej (exoccipital) w widoku od tyłu,
- pięć lub więcej kręgów krzyżowych (u dorosłych osobników - u młodych celofyzydów i Dilophosaurus są cztery),
- wklęsła proksymalna powierzchnia kości piszczelowej,
- skierowany dorsowentralnie, rozciągający się z proksymalnej powierzchni stawowej grzebień piszczelowy,
- najbardziej przyśrodkowa dystalna kość nadgarstka znacznie większa niż pozostałe dystalne kości nadgarstka,
- wystający wentralnie grzebień nadpanewkowy,
- obecność dużego dołu na wewnętrznej stronie zapanewkowej części kości biodrowej (dla przyczepu mięśnia caudifemoralis brevis),
- grzbietowa część kości biodrowej rozrośnięta dorsalnie (wyraźnie wyższa niż grzbietowa część obrzeży nadpanewkowej do styku kości łonowej i kulszowej),
- długi, rozciągnięty przed panewkę i kwadratowy przedni (przedpanewkowy) wyrostek kości biodrowej,
- wklęsły tylnoboczny brzeg dystalnej części piszczeli,
- wyraźny proksymodystalnie zwrócony grzebień na tylnej powierzchni dystalnej części piszczeli,
- spiczasta tylna odnoga dystalnej części IV kości stępu,
- kość skokowa zrośnięta z kością piętową,
- brak wyraźnie otoczonego i eliptycznego dołu za przednim wyrostkiem wstępując ym na tylnej powierzchnia kości skokowej,
- I kość śródstopia nieosiągająca proksymalnej powierzchni II kości i przyłączona do jej przyśrodkowej strony
Neoteropody dzieli się tradycyjnie rozłącznie na dwie grupy - Tetanurae i Ceratosauria (w których skład wchodzą celofyzoidy i ceratozauroidy). Jednak obecnie uważa się raczej, że celofyzoidy są bardziej bazalne od tetanurów i właściwych ceratozaurów (zob. też artykuł o ceratozaurach i poniżej). Neoteropody dzieli się więc na dwie nazwane grupy: celofyzoidy i awerostry (w skład których wchodzą tetanury i ceratozaury), między którymi plasuje się prawdopodobnie wiele form przejściowych.
Teropody przeważnie zaliczane do Coelophysoidea przez długi czas uważano za grupę monofiletyczną. Nowsze analizy (Yates, 2005; Smith i in., 2007; Nesbitt i in., 2009 i oparte na niej) wskazują, że nie tworzą one kladu a charakterystyczna budowa tych zwierząt to tylko 'celofyzoidowa faza' rozwoju teropodów (np. budowa przedniej części pyska z wcięciem między kością przedszczękową a szczękową, związana być może z elastycznością kości czaszki lub przystosowaniem do sprawnego manipulowania małą zdobyczą; podobną strkturę szczęki ma zauropodomorf Pampadromaeus). Wiele cech uważanych za unikalne dla nich mają już formy prymitywniejsze (Eoraptor i szczególnie kluczowa dla zrozumienia filogenezy wczesnych teropodów Tawa) a także te, które okazują się być bardziej zaawansowane (Zupaysaurus, Dilophosaurus). Analiza opublikowana w pracy Martineza i in. (2011), uwzględniająca Tawa i Eodromaeus, wspiera tę pierwszą hipotezę, lecz nie zawiera innych istotnych taksonów (jednakże rodzaj Tawa staje się bardziej bazalny przy wydłużeniu drzewka o jedną jednostkę, co powoduje, że celofyzoidy nie są monofiletyczne).
To samo dotyczy bliższych awerostrom dilofozaurydów (Dilophosauridae Madsen i Welles, 2000) - nowo rozpoznanej (lecz nie zdefiniowanej) grupy wyodrębnionej z celofyzoidów, co do której monofiletyczności także nie ma już pewności - Cryolophosaurus (wg analizy Smith i in, 2007 takson siostrzany Dilophosaurus) wydaje wg analizy Nesbitta i in. (2009) się bardziej bazalny od dilofozaura (to samo dotyczy innego teropoda uznanego przez Yatesa za przedstawiciela tej grupy - Zupaysaurus). Z awerostrami łączy je 5 cech czaszki: tylnobrzusznie wydłużony oczodół, kości szczękowe prawie równoległe do siebie patrząc od tyłu, zredukowana liczba zębów w kości szczękowej, językokształtny wyrostek na kościach ciemieniowych pokrywający guz nadpotyliczny i relatywnie głęboka kość kątowa górna. Także same dilofozaurydy zdiagnozowano na podstawie cech budowy czaszki: udziału wyrostka kości przedszczękowej w podłużnym i ostrym grzebieniu nosowym, przedłużeniu dołu przedoczodołowego w bocznowewnętrzną stronę kości nosowej i obecności nosowo-łzowego grzebienia (za Smith i in. 2007; cechy te mogą być szerszej rozpowszechnione; Smith i in. 2007 użyli pięciu cech związanych z budową grzebieni - po ich rewizji przez Brusattego i współpracowników (2010A) i odrzuceniu dwóch z nich klad ten nie został uzyskany). Jeśli taksony zaliczone przez Smitha i in. do tej grupy rzeczywiście tworzą klad, to są to teropody wczesnojurajskie (może też późnotriasowe), kosmopolityczne (znaleziono je w Afryce, Ameryce Północnej, Antarktydzie i Azji) i zadziwiająco podobne do siebie - wszystkie miały 6-7 m długości, smukłe, mocno wydłużone i niskie ciała, a na czaszkach fantazyjne ozdoby. Mniejsze lub większe grzebienie na czaszkach miały niemal wszystkie niecelofyzoidowe neoteropody (i może też celofyzoidy - "Megapnosaurus" kayentakatae; słabo rozwinięte miał Coelophysis), prócz tych, których czaszka została mocno zmodyfikowana (bezgrzebieniaste były Limusaurus, niektóre abelizaurydy i tyranozauroidy oraz przede wszystkim maniraptorokształtne celurozaury - wśród których niektóre znów wykształciły ozdoby).
Ewolucja: awerostry
We wczesnej jurze miejsce prymitywniejszych teropodów zajęły bardziej zaawansowane ceratozaury (Ceratosauria) i tetanury (Tetanurae), należące do Averostra. Ich nazwa oznacza dosłownie "ptasie dzioby". Przynależność do awerostrów można zdiagnozować na podstawie wielu cech, m. in. (za Smith i in., 2007):
- asymetryczne zęby kości przedszczękowej (pozbawione prostych stożków)
- brak otworu w kości kwadratowej,
- prezygapofyzje położone bocznie do kanału neuralnego w przednich kręgach szyjnych,
- łopatka nierozrośnięta znacznie na dystalnym końcu,
- wyraźny wewnętrzny "hak" na przedniej części kości biodrowej,
- szeroki, "skrzydełkowaty" mniejszy krętarz,
- zredukowana powierzchnia strzałkowa na kości skokowej,
- płaski wstępujący wyrostek skokowy.
Ceratozaury (Ceratosauria) to grupa teropodów znana głównie z kontynentów południowych. Bardziej zaawansowane z nich - abelizauroidy (Abelisauroidea) - dotrwały do końca kredy. Wyróżnia się wśród nich abelizaurydy (Abelisauridae) i noazaurydy (Noasauridae).
Takson siostrzany ceratozaurów to tetanury (Tetanurae), dzielące się na dwie główne grupy. Pierwszą są wymarłe w późnej kredzie megalozauroidy (Megalosauroidea), zawierające spinozaurydy (Spinosauridae) i megalozaurydy (zwane też torwozaurydami - Megalosauridae/Torvosauridae). Drugą żyjące do dziś aweteropody (Avetheropoda, inaczej neotetanury - Neotetanurae), do których należą celurozaury i wymarłe pod koniec kredy karnozaury (Carnosauria).
Celurozaury (Coelurosauria) były i są znacznie bardziej zróżnicowane niż ich należący do Carnosauria kuzyni. Prymitywnymi celurozaurami były tyranozauroidy (Tyrannosauroidea) i kompsognatydy (Compsognathidae). Bardziej zaawansowane od nich są maniraptorikształtne (Maniraptoriformes), dzielące się na dwie grupy - ornitomimozaury (Ornithomimosauria) i maniraptory (Maniraptora). Niepewną pozycję wśród nich mają alwarezzauroidy (Alvarezsauroidea). Maniraptory dzieliły się na kilka grup: terizinozaury (Therizinosauria), owiraptorozaury (Oviraptorosauria) i parawiale (Paraves). Wśród tych ostatnich wyróżnia się klad Eumaniraptora, do którego należą deinonychozaury (Deinonychosauria), dzialące się na dromeozaurydy (Dromaeosauridae) i troodontydy (Troodontidae). Do drugiej grupy eumaniraptorów zalicza się ptaki. Poza kompsognatydami, wszystkie wyróżnione wśród celurozaurów grupy dotrwały do końca mezozoiku. Wymieranie kredowe przetrwały zaś jedynie niektóre z tych należących do Avialae (geneza ptaków to obszerniejszy temat, który zostanie omówiony osobno).
Wymiary:
[5 cm (Mellisuga helenae)], ~43 cm (Parvicursor) < << 16-18 m (Spinosaurus)
Pierwsze teropody prawdopodobnie były małe (1-2 m długości), lecz niektóre bardzo szybko powiększyły rozmiary: (?Herrasaurus 4-6 m; Gojirasaurus >5 m; późniejszy Liliensternus >4-6 m). Z późnego triasu znane są też tropy i szczątki teropodów wielkości Allosaurus (Sander i in., 2011). Jurajskie niecelofyzoidowe neoteropody były już całkiem spore - osiągały ok. 6-7 m. Gigantyzm i prawdopodobnie zdolność do szybkiego powiększania rozmiarów ciała cechuje wiele grup teropodów. Największe z nich były prawdopodobnie bliskie osiągnięcia maksymalnych rozmiarów, jakie może mieć zwierzę dwunożne. Zdarzała się także miniaturyzacja - lecz było to zjawisko rzadsze; nieptasi przedstawiciele omawianej grupy (podobnie jak inne dinozaury) nigdy nie zbliżyli się nawet do małych rozmiarów osiąganych przez wiele ptaków, ssaków czy łuskonośnych.
Kiedy:
232-230 Ma -> dziś
najstarsze:
Eodromaeus - 232-230 Ma
najmłodsze: dzisiejsze ptaki
Teropody pojawiły najpóźniej 232-230 Ma (Furin i in., 2006), czyli w późnym (późny karnik wg skali Walkera i Geissmana, 2009) lub środkowym triasie (późny ladyn wg skali międzynarodowej z 2009). Z tego okresu z Ameryki Południowej znany jest najstarszy niewątpliwy teropod - Eodromaeus oraz kolejne dinozaury, które mogą należeć do tej grupy (herrerazaury oraz Eoraptor). Istniało już wtedy także 4-5 linii rozwojowych Sauropodomorpha (Chromogisaurus, Panphagia, Saturnalia, klad bardziej zaawansowanych od Guaibasauridae zauropodomorfów oraz Eoraptor, jeśli jest jednym z nich), co wskazuje na to, że teropody i ich najbliżsi krewni oddzielili się od siebie dużo wcześniej. Nieopublikowane badania (Dececchi i Larsson, 2007) wskazują, że pierwsze teropody pojawiły się już 5-7 milionów lat wcześniej, a więc ok. 239-235 Ma. Poza Ameryką Południową, domniemane szczątki najstarszych teropodów odkryto w Ameryce Północnej, Europie oraz w Indiach. Do jury przetrwali jedynie przedstawiciele Neotheropoda, formy bardziej bazalne, jak Daemonosaurus, wymarły wcześniej.
Gdzie:
Cały świat
Ponieważ teropody pojawiły się, gdy znane dziś kontynenty tworzyły jeden ląd, to - poza barierami środowiskowymi - nie było przeszkód w ich rozprzestrzenianiu się, lecz w triasie były rzadkie i mało zróżnicowane. W miarę rozpadu masy lądowej postępowała izolacja, lecz niektóre teropody prawdopodobnie zdołały w jakiś sposób dostać się na kontynenty oddzielone morzami (np. wędrując z wyspy do wyspy). Z drugiej strony niektóre grupy cechuje endemizm - lecz jest to często zjawisko trudne do zbadania.
Dowiedz się więcej:
- o zwierzętach, które mogły być teropodami:
(Agnosphitys), Alwalkeria (częściowo), "Beelemodon", "Capitalsaurus", (Chingkankousaurus), Eoraptor, Guaibasaurus, "Katsuyamasaurus", Protoavis, (Sinocoelurus), (Sinosaurus), (Velocipes)
- ? herrerazaurach:
??Chindesaurus, Herrerasaurus, ?Staurikosaurus, Sanjuansaurus
- bazalnych teropodach:
(?)Daemonosaurus, Eodromaeus, Tawa
- bazalnych, niecelofyzoidowych neoteropodach (w tym dilofozaurydach):
?Cryolophosaurus, ?Dilophosaurus, ?Dracovenator, ?Zupaysaurus
- i teropodach o niepewnej pozycji:
"Coelurosaurus", (Chienkosaurus), (Diplotomodon), (Embasaurus), "Futabasaurus", (Inosaurus), Labocania, ?"Merosaurus", ("Morosaurus" marchei), "Newtonsaurus", "Ngexisaurus", ("Ornithocheirus" hilsensis), "Saltriosaurus", (?)(Tanystrosuchus), (Teinurosaurus), (Thecocoelurus), (Wakinosaurus), (Walgettosuchus)

Rekonstrukcja Tawa hallae. Autor: Jorge Gonzalez. Rysunek pochodzi z tej strony.

Rekonstrukcje szkieletów bazalnych i prawdopodobnych teropodów. Od góry: A - herrerazaur Herrerasaurus ischigualastensis (rysunek pochodzi z tej strony), ?teropod B - Eoraptor lunensis (autor: Daniel Banchero; rysunek pochodzi z tej strony), C - bazalny teropod Eodromaeus murphi (autor: C. Abraczinskas; rysunek pochodzi z publikacji Martinez i in., 2011), bazalny teropod Tawa hallae (autor: Sterling Nesbitt; rysunek pochodzi z pracy Nesbitt i in., 2009), E - neoteropod (celofyzoid) Coelophysis bauri (autor: Gregory S. Paul; rysunek pochodzi z tej strony), F - neoteropod (celofyzoid lub bardziej zaawansowany neoteropod) Liliensternus liliensterni (autor: Gregory S. Paul; rysunek pochodzi z publikacji Paul, 1988), H - neoteropod (dilofozauryd lub ?celofyzoid) Dilophosaurus wetherilli (autor: Ville Sinkkonen; rysunek pochodzi z tej strony).

Kladogram z Yates, 2006 (na podstawie Rauhut, 2003 z rozbiciem Megapnosaurus na gatunki, połączeniem celurozaurów bardziej zaawansowanych od Proceratosaurus w jedną jednostkę, wyłączeniem Acrocanthosaurus z Carcharodontosauridae, dodaniem Dracovenator, Masiakasaurus, Dubreuillosaurus, Tugulusaurus i Zupaysaurus oraz 28 nowych cech i rewizją 5).

Kladogram ze Smith i in., 2007. Zob. też Mortimer, online.

Kladogram z Xu i in., 2009 (na podstawie Smith i in., 2007 z dodanymi 59 cechami z Carrano i Sampson, 2008 oraz dodatkowymi taksonami).

Kladogram z Nesbitt i in., 2009. Cyframi oznaczono klady: 72 - Theropoda, 60 - Herrerasauria, 69 - Neotheropoda, 61 - Coelophysoidea, 64 - Averostra, 63 - Tetanurae, 62 - Avetheropoda.

Kladogram z Knoll, 2010 (na podstawie skupionego na bazalnych zauropodomorfach badania Yatesa, 2007 ze zmianami na podstawie Smith i Pol, 2007 oraz dodanym Ignavusaurus).
Inna modyfikacja analizy (dodanie Chromogisaurus, Panphagia i nienazwanego herrerazaura MACN-PV 18649a oraz 15 cech) Smitha i Pola (2007) - dokonana przez Ezcurrę (2010) wskazała, że Agnosphitys i Guaibasaurus są zauropodomorfami, Chindesaurus ma niepewną pozycję wśród Eusaurischia, a do teropodów należy Eoraptor.

Kladogram z Cabreira i in., 2011 (na podstawie Ezcurra, 2010, na podstawie Yates, 2007).

Kladogram z Martinez, 2011 (na podstawie Sereno, 1999 z wieloma zmianami).

Skalibrowany stratygraficznie kladogram z Sues i in., 2011 (na podstawie Nesbitt i in., 2009 z dodanym Daemonosaurus i czterema nowymi cechami). "J Ther" to bardziej zaawansowane, jurajskie teropody.

Kladogramy z Langer i in., 2011 (na podstawie Nesbitt i in., 2009 z dodanym Guaibasaurus). Na lewo wersja niezmodyfikowana, na prawo z 6 nowymi cechami i usuniętymi nieistotnymi taksonami.

Kladogram z Nesbitt, 2011.

Skalibrowany stratygraficznie kladogram z Ezcurra i Brusatte, 2011 (na podstawie Nesbitt i in., 2009 z dodanymi Camposaurus i Megapnosaurus rhodesiensis oraz 24 nowymi cechami).

Kladogram z Tykoski, 2005 - konsensus Adamsa. W ścisłym konsensusie politomię tworzyły: celofyzydy (Coelophysis, Megapnosaurus, "M." kayentakatae, Segisaurus i nienazwany takson Shake-N-Bake), Eustreptospondylus u podstawy Tetanurae i Abelisaurus u podstawy Abelisauridae. Cienką linią zaznaczono pozycje w ścisłym konsensusie niekompletnych i potencjalnie niediagnostycznych taksonów, które dodawano do właściwej analizy osobno.
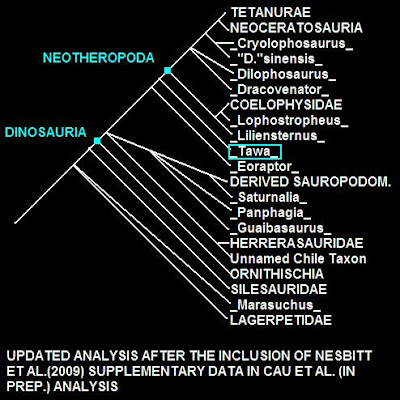
Kladogram z Cau, online 1. "Unnaned Chile Taxon" to nienazwany teropod z późnej jury Chile (Salgado i in., 2008).

Kladogram z Cau, online 2. Obrazek pochodzi z tej strony (kliknij na niego aby powiększyć).

Kladogram z Cau, online 3. Obrazek pochodzi z tej strony (kliknij na niego aby powiększyć).
Źródła:
Publikacje naukowe:
Agnolin, F.L., Ezcurra, M.D., Pais, D.F. & Salisbury, S.W. (2010) "A reappraisal of the Cretaceous non-avian dinosaur faunas from Australia and New Zealand: evidence for their Gondwanan affinities" Journal of Systematic Palaeontology 8, 257-300.
Alcober, O.A., Martinez, R.N. (2010) "A new herrerasaurid (Dinosauria, Saurischia) from the Upper Triassic Ischigualasto Formation of northwestern Argentina" ZooKeys, 63, 55-81. doi: 10.3897/zookeys.63.550
Balter, M. (2011) "Pint-Sized Predator Rattles The Dinosaur Family Tree" Science, 331(6014), 134. doi: 10.1126/science.331.6014.134
Barrett, P.M., Butler, R.J. & Nesbitt, S.J. (2011) "The roles of herbivory and omnivory in early dinosaur evolution" Earth and Environmental Science Transactions of the Royal Society of Edinburgh, 101, 383-396. doi: 10.1017/S1755691011020111
Benson, R.B.J. (2010) "The osteology of Magnosaurus nethercombensis (Dinosauria, Theropoda) from the Bajocian (Middle Jurassic) of the United Kingdom and a re-examination of the oldest records of tetanurans" Journal of Systematic Palaeontology 8, 131-146.
Brusatte, S.L., Benson, R.B.J., Currie, P.J. & Zhao, X.-J. (2010A) "The skull of Monolophosaurus jiangi (Dinosauria: Theropoda) and its implications for early theropod phylogeny and evolution" Zoological Journal of the Linnean Society, 158, 573-607.
Brusatte, S. L., Nesbitt, S. J., Irmis, R. B., Butler, R. J., Benton, M. J. & Norell, M. A. (2010B) "The origin and early radiation of dinosaurs" Earth-Science Reviews, 101, 68-100.
Cabreira, S.F., Schultz, C.L., Bittencourt, J.S., Soares, M.B., Fortier, D.C., Silva, L.R. & M.C. Langer (2011) "New stem-sauropodomorph (Dinosauria, Saurischia) from the Triassic of Brazil" Naturwissenschaften. doi: 10.1007/s00114-011-0858-0
Carrano, M.T. Hutchinson, J.R. & Sampson, S.D. (2005) "New information on Segisaurus halli, a small theropod dinosaur from the Early Jurassic of Arizona" Journal of Vertebrate Paleontology, 25, 835-849.
Ezcurra, M. D. (2010) "A new early dinosaur (Saurischia: Sauropodomorpha) from the Late Triassic of Argentina: a reassessment of dinosaur origin and phylogeny" Journal of Systematic Palaeontology, 8, 371-425.
Ezcurra, M.D. & Brusatte, S.L. (2011) "Taxonomic and phylogenetic reassessment of the early neotheropod dinosaur Camposaurus arizonensis from the Late Triassic of North America" Palaeontology, 54, 763-772. doi: 10.1111/j.1475-4983.2011.01069.x
Ezcurra, M.D. & Cuny, G. (2007) "The coelophysoid Lophostropheus airelensis, gen. nov.: a review of the systematics of "Liliensternus"airelensis from the Triassic-Jurassic boundary outcrops of Normandy (France)" Journal of Vertebrate Paleontology, 27, 73-86.
Ezcurra, M.D. & Novas, F.E. (2007) "Phylogenetic relationships of the Triassic theropod Zupaysaurus rougieri from NW Argentina" Historical Biology, 19, 35-72.
Knoll, F. (2010) "A primitive sauropodomorph from the upper Elliot Formation of Lesotho" Geological Magazine, 147, 814-829. doi:10.1017/S001675681000018X
Langer, M. C. (2004) "Basal Saurischia" [w:] Weishampel, D.B., Dodson, P. & Osmólska, H. "The Dinosauria" Berkeley and Los Angeles: University of California Press, 25-46.
Langer, M.C., Bittencourt, J.S. & Schultz, C.L. (2011) "A reassessment of the basal dinosaur Guaibasaurus candelariensis, from the Late Triassic Caturrita Formation of south Brazil" Earth and Environmental Science Transactions of the Royal Society of Edinburgh, 101, 301–332. doi: 10.1017/S175569101102007X
Langer, M.C., Ezcurra, M.D., Bittencourt, J.S. & Novas, F.E. (2009) "The origin and early evolution of dinosaurs" Biological Reviews, 84, 1-56.
Martinez, R.N., Sereno, P.C., Alcober, O.A., Colombi, C.E., Renne, P.R., Montañez, I.P. & Currie, B.S. (2011) "A basal dinosaur from the dawn of the dinosaur era in southwestern Pangaea" Science, 331, 206-210. doi: 10.1126/science.1198467
Nesbitt, S.J. (2011) "The early evolution of archosaurs : relationships and the origin of major clades" Bulletin of the American Museum of Natural History, 352, 1-292. doi: 10.1206/352.1
Nesbitt, S.J., Irmis, R.B. & Parker, W.G. (2007) "A critical re-evaluation of the Late Triassic dinosaur taxa of North America" Journal of Systematic Palaeontology, 5, 209-243.
Nesbitt, S.J., Smith, N.D., Irmis, R.B., Turner, A.H., Downs, A. & Norell, M.A. (2009) "A complete skeleton of a Late Triassic saurischian and the early evolution of dinosaurs" Science, 326, 1530-1533.
Nesbitt, S.J., Sidor, C.A., Irmis, R.B., Angielczyk, K.D., Smith, R.M.H. & Tsuji, L.M.A. (2010) "Ecologically distinct dinosaurian sister group shows early diversification of Ornithodira" Nature 464, 95-98.
Niedźwiedzki, G., Gorzelak, P. & Sulej, T. (2010) "Bite traces on dicynodont bones and the early evolution of large terrestrial predators" Lethaia, 44(1), 87–92. DOI: 10.1111/j.1502-3931.2010.00227.x
Salgado, L., De la Cruz, R., Suarez, M., Fernandez, M., Gasparini, Z., Palma-Heldt, S. & Fanning, M. (2008) "First Late Jurassic Dinosaur Bones from Chile" Journal of Vertebrate Paleontology, 28, 529-534.
Sander, P.M., Christian, A., Clauss, M., Fechner, R., Gee, C.T., Griebeler, E.-M., Gunga, H.-C., Hummel, J., Mallison, H., Perry, S.F., Preuschoft, H., Rauhut, O.W.M., Remes, K., Tütken, T., Wings, O. & Witzel, U. (2011) "Biology of the sauropod dinosaurs: the evolution of gigantism" Biological Reviews, 86(1), 117–155. doi: 10.1111/j.1469-185X.2010.00137.x
Sereno, P.C. (2007) "The phylogenetic relationships of early dinosaurs: a comparative report" Historical Biology, 19, 145-155.
Sereno, P.C. & Novas, F.E. (1992) "The complete skull and skeleton of an early dinosaur" Science, 258, 1137-1140.
Smith, N.D., Makovicky, P.J., Pol, D., Hammer, W.R. & Currie, P.J. (2007) "Osteology of Cryolophosaurus ellioti (Dinosauria: Theropoda) from the Early Jurassic of Antarctica and implications for early theropod evolution" Zoological Journal of the Linnean Society, 151, 377-421.
Sues, H.-D., Nesbitt, S.J., Berman, D.S. & Henrici, A.C. (2011) "A late-surviving basal theropod dinosaur from the latest Triassic of North America" Proceedings of the Royal Society B, 278, 1723, 3459-3464. doi:10.1098/rspb.2011.0410
Therrien, F. & Henderson, D.M. (2007) "My theropod is bigger than yours...or not: estimating body size from skull length in theropods" Journal of Vertebrate Paleontology, 27, 108-115.
Tykoski, R.S. & Rowe, T. (2004) "Ceratosauria" [w:] Weishampel, D.B., Dodson, P. & Osmólska, H. "The Dinosauria" Berkeley and Los Angeles: University of California Press, 151-168.
Wu, X., Currie, P.J., Dong, Z., Pan, S. & Wang, T. (2009) "A new theropod dinosaur from the Middle Jurassic of Lufeng, Yunnan, China" Acta Geologica Sinica 83(1), 9-24. doi:10.1111/j.1755-6724.2009.00002.x
Xu, X., Clark, J.M., Mo J., Choiniere, J., Forster, C.A., Erickson, G.M., Hone, D.W.E., Sullivan, C., Eberth, D.A., Nesbitt, S., Zhao, Q., Hernandez, R., Jia, C., Han, F. & Guo, Y. (2009) "A Jurassic ceratosaur from China helps clarify avian digital homologies" Nature, 459, 940-944.
Yates, A.M. (2006) "A new theropod dinosaur from the Early Jurassic of South Africa and its implications for the early evolution of theropods" Palaeontologia Africana, 41, 105-122.
Inne:
Dececchi, T. & Larsson, H. (2007) "Tempos and models of theropod evolution" Journal of Vertebrate Paleontology, 27, 67A [abstrakt]
Tykoski, R.S (2005) "Anatomy, Ontogeny, and Phylogeny of Coelophysoid Theropods" Dysertacja doktorska, University of Texas