


Jesteś w: Artykuły o teropodach
Tyranozauroidy (Tyrannosauroidea) Osborn, 1906 sensu Walker, 1964
Autorzy: Dawid Mika i Maciej Ziegler, korekta: Tomasz Sokołowski, Tomasz Skawiński, Tymoteusz Podwyszyński
Ostatnia aktualizacja: 21.11.2011 r.
Nomenklatura:
- Tyrannosauroidea (Osborn, 1906 sensu Walker, 1964)
(>Tyrannosaurus rex ~ Allosaurus fragilis v Ornithomimus velox v Deinonychus antirrhopus) (zmodyfikowano z Holtz, 2004) - Proceratosauridae (Averianov, Krasnolutskii i Ivantsov, 2010).
(>Proceratosaurus bradleyi ~ Tyrannosaurus rex v Allosaurus fragilis v Compsognathus longipes v Coelurus fragilis v Ornithomimus velox v Deinonychus antirrhopus) (zmodyfikowano z Rauhut, Milner i Moore-Fay, 2010)
Inna definicja to (<Proceratosaurus bradleyi & Kileskus aristotocus) (zmodyfikowano z Averianov, Krasnolutskii i Ivantsov, 2010). Na potrzeby artykułu będziemy używać tej pierwszej, ponieważ jest starsza i daje bardziej stabilny zasięg nazwy kladu. - Tyrannosauridae (Osborn, 1906).
(<Tyrannosaurus rex & Tarbosaurus bataar & Albertosaurus sarcophagus & Gorgosaurus libratus & Alectrosaurus olseni & Alioramus remotus & Daspletosaurus torosus) (Brochu, 2003)
W artykule przyjęto tę definicję, gdyż wcześniejsze (Sereno, 1998; Holtz, 2001) są problematyczne, ponieważ są oparte na taksonach o niejasnej pozycji filogenetycznej i ważności (Aublysodon, Nanotyrannus). Często używa się dla Tyrannosauridae definicji Holtza (2004), która zawęża zasięg nazwy do kladu Albertosaurinae+Tyrannosaurinae i pomija rodzaje o niepewnej pozycji filogenetycznej - Alioramus i szczególnie Alectrosaurus, który prawdopodobnie znajduje się poza Tyrannosauridae sensu Holtz (2004), podobnie jak Appalachiosaurus i Bistahieversor. - Albertosaurinae (Currie, Hurum i Sabath, 2003).
(>Albertosaurus sarcophagus ~ Tyrannosaurus rex) (zmodyfikowano z Currie i in., 2003) - Tyrannosaurinae (Osborn, 1906 sensu Matthew i Brown, 1922).
(>Tyrannosaurus rex ~ Albertosaurus sarcophagus) (zmodyfikowano z Currie i in., 2003)
Wcześniejsza definicja Sereno (1998; zmodyfikowana tutaj) to (>Tyrannosaurus rex ~ Albertosaurus sarcophagus v Daspletosaurus torosus v Gorgosaurus libratus). Na potrzeby artykułu będziemy używać późniejszej, ponieważ od 2003 roku powszechnie używa się nazwy Tyrannosaurinae w takim znaczeniu. - Inne (Cope, 1866 emmend. Brown, 1914 sensu Tatarinov, 1964). Prawidłowymi nazwami dla omawianej grupy, jak i dla Tyrannosauridae, mogą być wcześniej utworzone Deinodontoidea i Deinodontidae (Cope, 1866 emmend. Brown, 1914) często używane przed latami 50-tymi XX wieku. Cope, ustanawiając rodzinę Deinodontidae dla Deinodon, popełnił jednak błąd, pisząc Dinodontidae. Zostało to poprawione w 1914 r. przez Browna (już po utworzeniu Tyrannosauridae). Deinodon to obecnie nieważny (nomen dubium) rodzaj ustanowiony na podstawie zębów należących do bliżej nieokreślonego tyranozauryda. Nazwy Deinodontoidea i Deinodontidae dziś są praktycznie nieużywane i nie mają nadanej definicji filogenetycznej.
Podobnie Dryptosauridae (Marsh, 1890) - zależnie od pozycji filogenetycznej Dryptosaurus może być to prawidłowa nazwa dla grupy zaawansowanych tyranozaurydów (Tyrannosauridae). Nazwa Dryptosauridae jest dziś praktycznie nieużywana i nie ma definicji.
Charakterystyka:
-Co to gatunek?
-Tyranozaur.
-Matko Boska...
Michael Crichton ''Zaginiony Świat (The Lost World)
Tyrannosauroidea to klad drapieżnych, bazalnych celurozaurów. Nowe znaleziska i badania pozwoliły dobrze poznać tę grupę - można zaryzykować twierdzenie, że są najlepiej poznanymi wśród nieptasich teropodów.
Ewolucja i filogeneza
- Historia klasyfikacji. Od lat 20. XX wieku większość hipotez filogenetycznych plasowała tyranozauroidy jako potomków (Gilmore, 1920; Paul, 1988; Kurzanov, 1989) lub grupę siostrzaną (Molnar i in., 1990; Walker, 1964; Chatterjee, 1985; Bonaparte i in., 1990; Bonaparte, 1991) Carnosauria (choć niektóre współczesne analizy nadal na to wskazują - np. Göhlich i Chiappe, 2006 - także w wersji poprawionej przez Butlera i Upchurcha, 2007, jednak zapewne jest to spowodowane zbytnim ograniczeniem zakresu badania - analizowane były np. tylko dwa tyranozaurydy i dwa allozauroidy, a analiza dotyczyła bazalnych celurozaurów). W tych czasach teropody dzielono na dwie grupy: karnozaury i celurozaury, lecz często uważano je za bliżej niespokrewnione (Huene, 1932). Do pierwszej z nich wliczano wszystkie duże teropody, nie zważając na znaczące różnice w anatomii; podobnie do celurozaurów wliczano wszystkie niewielkie teropody. Huene (1920, 1926) ukuł termin Carnosauria dla monofiletycznego taksonu złożonego z Allosaurus i "megalozaurów" (=dużych europejskich teropodów). Uznał on tyranozauroidy za celurozaury bliskie ornitomimozaurom na podstawie zaawansowanej budowy stopy (jak się okazało później, u najbardziej bazalnych tyranozauroidów stopa ma prymitywną budowę). Podobnie uważali Matthew i Brown (1922) oraz Bakker (1977), Bakker i in. (1988). Gregory S. Paul w swojej słynnej książce pt. "Predatory Dinosaurs of the World" uznał za przodków tyranozauroidów allozaurydy, tymczasem Sankar Chatterjee "rauizuchy"! Analizy filogenetyczne Thulborna (1984), Novasa (1991), Pereza-Moreno i in. (1993; 1994), Holtza (1994), Sereno (1994) oraz późniejsze wykazały, że tyranozauroidy należą do celurozaurów. Badanie tego pierwszego wykazało, że omawiana grupa jest bliższa dzisiejszym ptakom niż Archaeopteryx (!). Obecnie uważa się, że Tyrannosauroidea to grupa bardzo bazalnych przedstawicieli Coelurosauria.
- Ewolucja. Tyranozauroidy zróżnicowały się już w środkowej jurze - w batonie (168-164 Ma). Jak wynika z najobszerniejszej dotąd analizy, jednak właściwie nietestującej przynależności do Tyrannosauroidea włączonych taksonów (Brusatte i in., 2010B), istniało już wtedy pięć linii rozwojowych: najbardziej bazalne mają być proceratozaurydy, natomiast pozostałe tworzą linię wiodącą do wielkich, słynnych tyranozaurydów. Sereno i in. (2009) podzielili ewolucję Tyrannosauroidea na trzy etapy, których zmodyfikowaną wersję prezentujemy poniżej. Kluczowy dla takiej gradacji jest Raptorex, który może być bardzo młodym, bardziej zaawansowanym tyranozauroidem. Ponieważ nie zostały podważone wyniki oględzin skały, w których zachował się okaz raptoreksa, które zdaniem Sereno i in. wskazują na wczesnokredowe warstwy Lujiatun, tymczasowo zachowujemy podział Sereno i in.:
1) Proceratosauridae. Najstarszymi znanymi tyranozauroidami są Kileskus (baton, 168-164 Ma) i prawdopodobnie też Proceratosaurus (późny baton, 166-164 Ma); młodszy jest późnojurajski Guanlong (pierwsza połowa oksfordu, 161-158 Ma). Wszystkie trzy należą być może do Proceratosauridae, jednak mogą one nie tworzyć kladu - sam Proceratosaurus (a wraz z nim także takson Proceratosauridae) może być kompsognatydem albo ornitomimozaurem (Choiniere i in., 2010). Teropody te łączą cechy czaszki (niektóre obecne też u innych teropodów): (1) krótka wewnętrzna krawędź kości przedszczękowej, (2) głębokość dołu przedoczodołowego dużo większa niż głębokość kości szczękowej pod wewnętrzną krawędzią tego dołu, (3) wstępujący wyrostek kości szczękowej odrastający z jej przedniego brzegu, (4) duże otwory nosowe oraz nieznane u Kileskus (5) grzebień na kości nosowej i (6) głęboki i ostry nerwo-naczyniowy rowek na kości zębowej; kolejną cechą diagnostyczną mogą być wydłużone tylne kręgi ogonowe z głębokimi, wzdłużnymi rowkami na grzbietowej stronie łuku kręgowego i wewnętrznej stronie trzonów (Rauhut i in., 2010; Averianov i in., 2010).
Sinotyrannus - duży (9-10 m długości i metrowa czaszka, ale zob poniżej - w "Wymiary") tyranozauroid z wczesnej kredy (122-112 Ma) Chin został początkowo uznany za zaawansowanego tyranozauroida - prawdopodobnie tyranozauryda (o których za chwilę). Jednak Ji in. (2009) podali - prócz wielkości - właściwie tylko jedną cechę czaszki upodabniającą go do przedstawicieli Tyrannosauridae - konkretnie Tyrannosaurinae (druga z nich - wyraźny pionowy grzebień na kości biodrowej jest obecny też u innych teropodów np. Aviatyrannis, Iliosuchus). Wskazali za to aż pięć różnic. Jak piszą Brusatte i współpracownicy (2010A), sinotyran ma przynajmniej niektóre (3-5) synapomorfie Proceratosauridae oraz jest podobny do proceratozaura i guanlonga w jeszcze jednym aspekcie - główne otworki nerwowo-naczyniowe kości zębowej umieszczone są w ostrym rowku. W analizie Awerianowa i in. także wczesnokredowy Dilong jest proceratozaurydem, jednak jedynym niebazalnym tyranozauroidem jest w niej Tyrannosaurus rex, co stawia pod dużym znakiem zapytania tę topologię.
Inne formy bazalne. Wczesne tyranozauroidy miały długie, praktycznie trójpalczaste kończyny przednie (u Guanlong obecna jest 4 kość śródręcza). Przyczyną redukcji kończyn przednich bardziej zaawansowanych tyranozauroidów i jednoczesne zwiększenie się i wzmacnianie szczęk był zapewne sposób polowania - mianowicie chwytanie ofiar szczękami, a nie kończynami przednimi. Ten sposób polowania utorował drogę innym cechom, które umożliwiły maksymalne zwiększenie siły szczęk - najbardziej zaawansowane tyranozaurydy - jak tyranozaur - miały największą siłę zacisku szczęk ze wszystkich teropodów. W późnej kredzie, obok słynnych, gigantycznych form, mogły żyć średniej wielkości prymitywne tyranozauroidy, znane z bardziej fragmentarycznych skamielin, jak pochodzący z mastrychtu Bagaraatan, który może być ornitomimozaurem.
2) Druga faza rozwoju tyranozauroidów jest obecnie reprezentowana przez Raptorex (ale zob. opis) i cechuje się bardziej widocznymi zmianami - powiększeniem czaszki, jej pneumatyzacją, zwiększeniem siły szczęk, dalszym zróżnicowaniem (z przodu szczęk małe i słabo zakrzywione, dalej położone coraz bardziej zakrzywione, największe pośrodku) i zmniejszeniem liczby zębów, skarłowaceniem i zmianą budowy kończyn przednich, wydłużeniem kończyn tylnych i powstaniem arctometatarsus (którego miał prawdopodobnie także Dryptosaurus). Tyranozauroidy te miały silnie rozrośnięte "nogi" i miednicę, co wskazuje na silnie mięśnie.
Australijski tyranozauroid (NMV P186046 - Benson i in., 2010 A) był nawet jeszcze bardziej zaawansowany, na co wskazuje wyraźny guzek przypominający kryzę i rozszerzona z przodu dystalna ekspansja kości łonowej (niepublikowana analiza Cau wskazuje jednak, że był on bardziej bazalny, ale bliższy tyranozaurydom niż Eotyrannus).
3) Tyrannosauridae. Do tej grupy należą najbardziej zaawansowane tyranozauroidy. Najstarsi pewni przedstawiciele Tyrannosauridae pochodzą z początków kampanu (ok. 83 Ma). Cechy charakteryzujące tyranozaurydy są trudne do stwierdzenia z powodu niepewnej pozycji filogenetycznej rodzajów takich jak Appalachiosaurus, Alectrosaurus i Bistahieversor, które są bardzo podobne do siebie - szczególnie w szkielecie pozaczaszkowym. Są one zapewne bardziej zaawansowane niż Raptorex, który wygląda już jak miniaturowy przedstawiciel Tyrannosauridae - początkowo wzięto go nawet za młodego tarbozaura. Różni się on od nich jedynie gładką powierzchnią u góry kości łonowej, gorzej rozwiniętym arctometatarsus, dłuższym trzonem kości ramiennej - między grzbietem deltapektoralnym a dystalnymi kłykciami, bocznie spłaszczonymi zębami (w przeciwieństwie do niemal cylindrycznych w przekroju zębów dorosłych tyranozaurydów, można więc wyjaśnić tę różnicę niedojrzałością holotypu raptoreksa) oraz cechami związanymi prawdopodobnie z wielkością tych teropodów: brakiem szypuły węchowej i niepodzielonym kresomózgowiem, znacznie - 1,18 razy - dłuższą piszczelą w porównaniu do kości udowej, powiększonym dołem przedoczodołowym, kostnymi kołnierzami w oczodole, bocznoskroniowymi otworami, wzniesieniem potylicy, wyrostkami na kręgach szyjnych dla przyczepu mięśni oraz skróceniem kręgosłupa. Gigantyzm tyranozaurydów na końcu kredy i prawdopodobnie powiązane znaczne zmiany w budowie ich mózgów w tym czasie najprawdopodobniej były spowodowane zwolnieniem niszy ekologicznych zajmowanych dużych mięsożerców - karnozaurów, choć do wyparcia innych tetanurów przynajmniej częściowo doszło już wcześniej (np. w Azji w apcie - ok. 122-112 Ma - dominował Sinotyrannus, lecz późniejsze karnozaury - Shaochilong i Chilantaisaurus - przynajmniej lokalnie stały na szczycie łańcucha pokarmowego). Panowanie tyranozaurydów zakończyło wielkie wymieranie kredowe, a sam zasięg był ograniczony.
Tyrannosauridae dzielą się na:
- Albertosaurinae, których przedstawiciele byli generalnie smuklejsi, z niższymi czaszkami oraz proporcjonalnie dłuższymi kośćmi podudzia (cechy najpewniej plezjomorficzne; Alioramus także był lekko zbudowany i miał niską czaszkę). Występowanie dwóch niewątpliwych albertozaurynów ograniczało się do północnej części Ameryki Północnej (wtedy przedzielonej na dwie części morzem). Nie dotrwały one do końca ery mezozoicznej, ustępując miejsca tyranozaurynom, które występowały zarówno w Azji, jak i w Ameryce Północnej.
- Tyrannosaurinae, cechujące się wypukłą grzbietową powierzchnią kości łuskowej, której sklepienie było przedziurawione pneumatycznym otworem, kwadratową lub rozdwojoną końcówką wyrostka jarzmowego kości kwadratowo-jarzmowej, szerokimi tylnie kośćmi przedczołowymi i wysokim grzebieniem sagitalnym na kości czołowej (za Carr i Williamson, 2010).
Karol Sabath i Jorn H. Hurum przebadali biomechanikę czaszek Tarbosaurus i Alioramus i Tyrannosaurus (żyjącego w Ameryce Północnej). U tarbozaura podczas gryzienia siła była przekazywana od kości szczękowej do otaczających je kości, tymczasem u tyranozaura od kości szczękowej do zrośniętych kości nosowych, a następnie do łzowych przez coś w rodzaju podpórek lub wsporników kostnych. Tarbozaur ich nie posiadał, ale miał mocno zbudowany występ kości szczękowej, mieszczący się w osłonie z kości łzowej. Ten sam wyrostek u amerykańskich tyranozaurydów, w tym tyranozaura, był zaledwie cienką płytką. Równocześnie mocne połączenie między kością łzową a kośćmi czołowymi i przedczołowymi świadczy o jej mniejszej elastyczności. Badanie to wykazało, że najbliższym krewnym tarbozaura jest alioram, a północnoamerykańscy i azjatyccy przedstawiciele Tyrannosaurinae ewoluowali niezależnie od siebie, choć nowe dane (opis drugiego gatunku aliorama - Brusatte i in., 2009) wskazują na to, że oba azjatyckie rodzaje nie tworzą kladu. - Pokrewieństwo między tyranozauroidami. Niektóre poglądy na temat filogenezy tyranozauroidów są w wielu analizach podobne, istnieją jednak znaczące różnice. Przykładowo w badaniu Xu i in. (2006, na podstawie Rauhut, 2003) guanlong i stokesozaur tworzą politomię z kladem tworzonym przez dilonga i tyranozaurydy (podobny wynik uzyskano w dalszej modyfikacji analizy Rauhuta z 2010 r.). Niepublikowane badanie Miyashity i Curriego (2009) wykazało natomiast, że to Guanlong jest bardziej zaawansowany od Dilong. W tej analizie Itemirus jest najbardziej bazalnym tyranozauroidem a Appalachiosaurus jest bardzo bliski tyranozaurydom.
Tyranozauroidy to grupa będąca obiektem wielu badań, także dotyczących pokrewieństwa - wyniki kolejnych analiz prezentujemy jako ryciny (zob. też kladogramy w artykule o Coelurosauria: Senter, 2007; 2010; Li i in., 2010). - Tyranozauroidy a inne bazalne celurozaury. Wiele bazalnych celurozaurów o niejasnej klasyfikacji, a także zaliczanych tradycyjnie do kompsognatydów, może okazać się tyranozauroidami. Dotyczy to: Calamosaurus, *Calamospondylus, *Embasaurus, Itemirus, *Mirischia, Santanaraptor i *Xinjiangovenator (oznaczone gwiazdką uznano za tyranozauroidy w niepublikowanych badaniach). Dla przykładu Calamosaurus wykazuje duże podobieństwo do dilonga, co sugeruje, że był jego bliskim krewniakiem (Naish i Martill, 2007). W jednej z analiz (Zanno i in., 2009) teropody zaliczane powszechnie do Compsognathidae należą do Tyrannosauroidea, jednak jedynym poza tyranozaurydami tyranozauroidem włączonym do analizy jest dilong, co zapewne wpłynęło na wynik. Taki sam wynik uzyskano w innej analizie ograniczonej do relacji między tyranozauroidami (Zanno i Makovicky, 2011 - na podstawie Sereno i in., 2009). Czasem uznaje się za tyranozauroidy takie teropody jak Bagaraatan (ornitomimozaur?), Iliosuchus i Labocania. Coelurus i Tanycolagreus mogą tworzyć klad najbardziej bazalnych tyranozauroidów (Senter, 2007; 2010).
Dilong miał cechy kompsognatydów (np. budowa miednicy) i w niektórych analizach (Turner i in., 2007; Lee i Worthy, 2011) okazał się być bliższy ptakom. Proceratosaurus może należeć do Tyrannosauroidea, ale może też być bazalnym kompsognatydem albo nawet ornitomimozaurem. Analiza Lee i Worthyego (2011) wykazała, że Dilong i Eotyrannus przedstawicielami nienazwanego kladu siostrzanego do Tyrannosauroidea, który zawiera Maniraptoriformes oraz wiele bardziej bazalnych od maniraptorokształtnych taksonów. Po raz pierwszy taki wynik uzyskano w nieopublikowanych badaniach Mortimera. U niego poza Dilong, Eotyrannus i Proceratosaurus dotyczy to Guanlong i Kileskus.
Generalnie bazalne celurozaury są do siebie bardzo podobne, a gdy weźmie się pod uwagę fragmentaryczność większości z tych taksonów, to nie powinny dziwić takie rozbieżności w klasyfikacji powyższych rodzajów. Z pewnością potrzebna jest zakrojona na szeroką skalę analiza pokrewieństwa domniemanych tyranozauroidów, zawierająca wiele bazalnych celurozaurów.
Wczesne tyranozauroidy i dromeozaurydy miały bardzo podobne zęby. Jeden ze znanych z zębów i szczęk eumaniraptorów (prawdopodobnie dromeozauryd) - Richardoestesia okazał się w nieopublikowanej analizie Cau (online) bazalnym tyranozauroidem.
Ewolucja i budowa
- Czaszka. U wczesnych tyranozauroidów były - jak u innych celurozaurów - niskie i lekko zbudowane. U najbardziej zaawansowanych tyranozauroidów stały się one wysokie i masywne. Kości przedszczękowe były nadzwyczajnie wysokie i krótkie (4-6% długości czaszki), co może być cechą także innych celurozaurów (Tanycolagreus), ze zbliżonymi do siebie wyrostkami nosowymi, na których podstawie znajdował się głęboki otwór oraz zwróconymi ku górze wyrostkami szczękowymi. Okno szczękowe stykało się z jamą szczękową w widoku przyśrodkowym. Tyranozauroidy posiadały też mocne (z wyjątkiem Xiongguanlong), łukowate kości nosowe (kolejna konwergencja z abelizaurydami). Wystający grzbiet kości kątowej ciągnie się aż do stawu żuchwy. Tyranozauroidy to stosunkowo prymitywne celurozaury, z bardziej "gadzim" niż "ptasim" i mniejszym niż u nich mózgiem (który u dorosłych nie wypełniał całej przestrzeni mózgoczaszki). Współczynnik encefalizacji (EQ) wynosił od 2 do 2,4, był więc pośredni między bardziej bazalnymi teropodami a tymi bliższymi ptakom. Cechują się także kością kwadratowo-jarzmową posiadającą wybrzuszenie i wygiętą dolną krawędź, zwężoną podstawą czaszki, spneumatyzowaną kością jarzmową (obecne też u wielu karnozaurów) z pomarszczonym brzusznym zgrubieniem jej pod oczodołem. U większości tyranozaurydów czaszka stała się dość wysoka a pysk głęboki, choć może być to już cechą Raptorex. Niektóre z jej kości - np. nosowe - stały się grubsze i połączyły się dla zwiększenia jej siły i wytrzymałości. Wzmocnienie góry czaszki, która jest silnie spneumatyzowana (podobnie jak żuchwa) cechuje tyranozaurydy i raptoreksa. Z drugiej strony u Alioramus (który może nie należeć do Tyrannosauridae) czaszka była bardzo długa i nie tak masywna. Połączone kości ciemieniowe miały strzałkowaty wyrostek grzebieniowaty, który znajdował się na górze czaszki. Bardziej zaawansowane tyranozauroidy cechują się proporcjonalnie wielkimi czaszkami - osiągającymi u Raptorex, Xiongguanlong i tyranozaurydów ok. 40 % długości tułowia (u innych teropodów, prócz długopyskich spinozaurydów, było to mniej niż 30 %). Największa znaleziona czaszka tyranozaura ma ponad 1,5 m długości. Niektóre z nich (Xiongguanlong, Alioramus) miały bardzo niskie czaszki, których długość przewyższała wysokość w okolicy okna przedoczodołowego pięciokrotnie, co jest cechą także spinozaurydów i unenlaginów (Austroraptor i Buitreraptor). Czaszki innych były wyższe i krótsze, a bardzo skróconą miał Teratophoneus. Inne cechy zaawansowane łączące raptoreksa i tyranozaurydy to szew między kością łzową i czołową w okolicy oczodołu, spneumatyzowane kości łzowe i łuskowe oraz otwór w kości kątowej górnej. W ewolucji tyranozauroidów zaobserwowano też zwiększanie mięśni zamykających szczęki, postępujące różnicowanie zębów z przodu górnej szczęki (były one mniejsze od dalszych i miały w przekroju kształt litery "D", co jest prawdopodobnie synapomorfią Coelurosauria; obecne m.in. u bazalnego celurozaura Zuolong, ornitomimozaura Pelecanimimus, karnozaura Allosaurus i dromeozauryda Deinonychus) i zwiększanie roli węchu. Zęby przedszczękowe służyły to zdrapywania a położone dalej do cięcia i rozrywania. Z biegiem czasu czaszki większości zaawansowanych tyranozauroidów (bez Alioramus) stały się mocne i przystosowane do przegryzania kości - zęby stały się kołkowate, styki kości pozrastały się lub zaryglowały, żuchwa stała się sztywna z ryglującym spojeniem.
- Kończyny przednie. Początkowo wyposażone w trójpalczaste kończyny, w biegu ewolucji utraciły trzeci palec kończyn przednich, które znacząco się zredukowały. U bazalnego tyranozauroida - Guanlong kończyny przednie mają długość równą mniej więcej 60% długości kończyn tylnych. Stosunkowo krótkie, trójpalczaste kończyny przednie występują u bardziej zaawansowanego Eotyrannus. Zaawansowane formy miały już bardzo krótkie ''rączki''. Szczytem redukcji kończyn przednich u tyranozauroidów są te należące do Tarbosaurus - długość jego kości ramieniowej wynosiła jedną czwartą długości kości udowej (u Raptorex - 29%, co może wynikać z jego młodego wieku). Tak jak u innych tyranozaurydów były one dwupalczaste, choć niektóre okazy Tyrannosaurus zdradzają obecność szczątkowego trzeciego palca. Prawdopodobnie już dryptozaur miał tylko dwa palce. Wydaje się, że nie były to narządy szczątkowe - były silne i z ograniczonym zakresem ruchów, co wskazuje na to, że były przystosowane do łapania zdobyczy. Nieopublikowane dane (Williams i in., 2010) wskazują, że łapy młodych tyranozaurydów, a przynajmniej samego tyranozaura, były proporcjonalnie znacznie dłuższe i smuklejsze niż dorosłych i były uzbrojone w ogromne pazury. Prawdopodobnie najpierw zmniejszyły się ramiona tyranozauroidów a dopiero później dłonie, na co wskazuje duża dłoń dryptozaura, lecz znacznie skrócone ramię (długości pośredniej między bazalnymi tyranozauroidami a zaawansowanymi - bliskimi tyranozaurydom).
- Kończyny tylne. Tyranozaudoidy, jak na teropody przystało, miały cztery palce, z których pierwszy nie dotykały gruntu podczas poruszania się. Były one w stosunku do długości ciała długie i - jak u szybko biegających zwierząt - miały wydłużoną piszczel i kości śródstopia. U młodych albertozaurów były one proporcjonalnie tak długie jak u ornitomimozaurów. Proporcje długości poszczególnych partii "nóg" zmieniają się u najbardziej znanego tyranozauroida - tyranozaura. Świadczy to o tym, że dorośli przedstawicieli tego rodzaju byli raczej chodziarzami niż biegaczami. Teropody te miały również wklęsłe nacięcie przy górnym końcu kości biodrowej (która jest bardzo długa, szczególnie u stokesozaura i eotyrana), poszerzającą się ku górze panewkę stawu biodrowego, wielkie dystalne rozszerzenie na końcu kości łonowej, podniesioną głowę kości udowej i ostro zakończoną nasadę kości strzałkowej.
- Ozdoby. U guanlonga na czaszce znajduje się pojedynczy kostny grzebień. Jest on podtrzymywany przez kości nosowe i biegnie od początku do końca czaszki. W grzebieniu guanlonga znajdowały się puste przestrzenie wypełnione za życia workami powietrznymi. Podobne grzebienie miały prawdopodobnie także proceratozaur i sinotyran. Mniej rzucające się w oczy wypukłości biegną wzdłuż boków czaszki dilonga. Jednak w odróżnieniu od tych występujących u guanlonga są one przytrzymywane także przez kości łzowe. Bardziej zaawansowany tyranozauroid - alioram - ma 3-6 (zależnie od gatunku) niewielkich kościstych guzów na kościach nosowych. Są one obecne, lecz niższe u Daspletosaurus, Appalachiosaurus, Gorgosaurus i Tarbosaurus. Chropowata powierzchnia góry czaszki zaawansowanych tyranozauroidów wskazuje na występowanie okrywy rogowej.
U Albertosaurus, Alioramus, Appalachiosaurus, Bistahieversor, Gorgosaurus i Daspletosaurus występuje niewielka, rogopodobna struktura nad okiem - na kościach łzowych. Owa struktura nie występuje u tarbozaura i tyranozaura, jednak posiadają one półksiężycowy grzebień nad każdym okiem wystający z kości nadoczodołowej. Owe ozdoby mogły służyć do rozpoznawania własnego gatunku lub stada albo wabienia samic. W przypadku grzebieni proceratozaurydów zasugerowano redukowanie nacisków na kości czaszki podczas gryzienia. - Prapióra. Najprawdopodobniej tyranozauroidy były opierzone, nie mamy jednak co do tego absolutnej pewności. Należałoby przyjąć, że były pokryte tylko łuskami, gdyby zostały spełnione łącznie następujące przesłanki: (1) dinozaury nie były pokryte prapiórami od samego początku (moim zdaniem [MZ] nieznacznie przeważa hipoteza, że były), (2) karnozaur Concavenator nie miał piór (co jest wysoce prawdopodobne - zob. opis), (3) opierzony Dilong nie jest tyranozauroidem (co jest możliwe: mimo że większość badań, szczególnie tych, które miały za zadanie zbadanie relacji między bazalnymi celurozaurami, wskazuje, że Dilong to tyranozauroid, to jednak w niektórych analizach - jak Turnera i in., 2007 oraz na niej opartych, np. Csiki i in., 2010, a także Turner i in., 2011 - Dilong okazał się bliższy ptakom niż tyranozauroidy; jednak często jedynymi innymi włączonymi do badania tyranozauroidami były tyranozaurydy, co zapewne wpłynęło na wynik; podobny wynik miało też badanie Lee i Worthy, 2011), (4) kompsognatydy (Juravenator, Sinocalliopteryx, Sinosauropteryx) są bliższe ptakom niż tyranozauroidy (na co wskazuje większość analiz) (5) odnaleziona w Chinach, pokryta prapiórami tylna część ogona zaliczona tymczasowo do Tyrannosauroidea przez Xu i in. (2010) nie należy do przedstawiciela omawianej grupy i wreszcie (6) pierzasty Juravenator jest celurozaurem bardziej zaawansowanym niż tyranozauroidy a nie jest bazalnym tetanurem - konkretniej megalozauroidem (na co wskazują wszystkie opublikowane badania, jednak niepublikowane dane Rauhuta i Fotha podobno wskazują, że Juravenator to bazalny megalozauroid, co oznacza, że niemal wszystkie znane tetanury, także tyranozauroidy, musiały być pierzaste - zob.komentarz na DML).
- Inne. Podobnie jak wiele neoteropodów, także tyranozauroidy miały szyję wygiętą w kształt litery "S" oraz długi ogon. Tyranozauroidy cechują się większym stopniem spneumatyzowania szkieletu (kręgosłupa i czaszki) w porównaniu do innych teropodów, co przyjęło najbardziej ekstremalne rozmiary u Alioramus. Prawdopodobnie nie było to związane z dążeniem do zmniejszenia masy zwierzęcia, gdyż stopień rozwinięcia tych struktur był większy u młodych tarbozaurów a mniejszy u starszych i większych.
Odżywianie i zmysły
Kwestią kontrowersyjną (aczkolwiek niesłusznie) jest strategia pokarmowa tyranozaurydów. Prof. Jack Horner, znany z opisania majazaurów (ornitopodów znanych z opieki rodzicielskiej) uważa, że tyranozaur i ogólnie tyranozaurydy były padlinożercami, których ewolucja była ukierunkowana w tę stronę. Jego poglądy zostały rozsławione przez film popularnonaukowy "Dolina tyranozaura" (Valley of T.rex) emitowany w Polsce na kanałach Discovery. Jako dowody na padlinożerność tyranozaura i innych tyranozaurydów przedstawia on:
- Tyranozaur był jego zdaniem zbyt wolny, aby dogonić żyjących w jego środowisku roślinożerców, takich jak hadrozaurydy.
- Silnie zredukowane kończyny przednie tyranozaurydów nie mogły służyć do chwytania ofiary.
- W czaszce tyranozaura bardzo dobrze rozwinięte są struktury odpowiedzialne za węch, a za wzrok małe. Podobne proporcje występują u współczesnych sępów i innych padlinożerców (dziś wiadomo, że to nieprawda - Witmer i Ridgely, 2009 - zob. poniżej).
- Horner uważa, że ze względu na przypuszczalny kiepski wzrok tyranozaur nie mógł polować w nocy.
- Potężne szczęki nie muszą służyć do polowania, lecz do miażdżenia kości padliny i wydobywania z nich pożywnego szpiku.
- W ciągu ewolucji tyranozaurydów dochodziło do zwiększania się siły nacisku szczęk, co jego zdaniem jest adaptacją do padlinożerności, gdyż to głównie oni potrzebują mocnych szczęk. Hieny (które jak się okazuje, często polują) mają większą siłę szczęk niż lwy (które czasami pożywiają się padliną).
- W ciągu ewolucji tyranozauroidów dochodziło do powiększenia się rozmiarów ciała. Im większe zwierzę, tym ma większe szanse na odpędzenie prawdziwych łowców od padliny, a tym samym pożywienie się i przetrwanie.
Podsumowując - Horner uważa tyranozaura za wielkiego ścierwojada ("przerośniętego sępa"), niezdolnego do upolowania jakiejkolwiek zdobyczy.
Jednak w jego rozumowaniu istnieją rażące błędy. Dobry węch nie musi świadczyć o padlinożerności - współczesne wilki mają dobry węch, a często polują. Tak samo obdarzone świetnym węchem hieny żyją głównie z polowań. Wiązanie dobrego węchu ze zwyczajami żywieniowymi także nie jest często trafne - mógł on służyć do innego rodzaju aktywności, np. lokalizowania innych osobników swego gatunku albo polowania przy słabym świetle. Jak wykazali Witmer i Ridgely (2009), to, co wcześniej brano za opuszki węchowe, to w rzeczywistości głównie okolica węchowa jamy nosowej. Część mózgu odpowiedzialna za węch była średniej wielkości, choć większa niż u większości teropodów (prócz dromeozaurydów), a u CMNH 7541 (holotyp Nanotyrannnus) była jeszcze bardziej powiększona. Tyranozaur (oraz nanotyran, jeśli są odrębne) miały bardzo dobry węch, choć nie aż tak, jak sądzono wcześniej. Dilong miał dużo gorszy węch niż tyranozaurydy. Oczodoły tyranozaura były ustawione pod kątem zapewniającym widzenie obuoczne - i to lepsze od jastrzębia. Widział stereoskopowo, co zapewniało dokładną ocenę odległości. W ciągu milionów lat oczy tyranozaurydów ustawiały się pod kątem zapewniającym coraz lepsze widzenie. Trudno wytłumaczyć czemu takie zmiany zaszły u potencjalnych padlinożerców. Współcześnie z obuocznego widzenia korzystają drapieżniki (w celu dokładniej oceny odległości między sobą i ofiarą) oraz naczelne (co umożliwia im dokładną ocenę odległości między gałęziami w celu ich przeskoczenia). Dla kontrastu tarbozaur miał oczy ustawione na boki i nie mógł widzieć przez to obuocznie. Tak więc paradoksalnie oczy drapieżnego tarbozaura były tak położone jak oczy współczesnych ssaków roślinożernym, którym takie położenie umożliwia sprawniejsze wypatrzenie wroga.
Zresztą, nawet jeśli przypuścimy, że tyranozaur miał słaby wzrok (czemu przeczą dowody) i tak był on z pewnością wystarczający do wypatrzenia wielkich roślinożerców - jak hadrozaurydy czy ceratopsy. Horner uważa, że niewielkie proporcjonalnie oczy tyranozaura są dowodem na padlinożerność, jednak i tu mija się z prawdą. Proporcjonalnie oczy tego dinozaura w porównaniu z oczami innych teropodów są średniej wielkości, a istnieją teropody o proporcjonalnie mniejszych oczach - jak Giganotosaurus czy Dilophosaurus. Ogólnie oczy tyranozaura były bardzo duże i mogły osiągać do 14 cm średnicy (Stevens, 2006).
Tyranozaurydy prawdopodobnie umiały szybko skupiać wzrok, co jest istotne przy szybkich ruchach. Snively i Russell (2007) wykazali, że T. rex miał szyję przystosowaną do gwałtownych ruchów bocznych, co w połączeniu z dobrym wzrokiem było zapewne przystosowaniem do atakowania innych zwierząt.
Także słuch tyranozaurydów był dobrze rozwinięty, szczególnie jeśli chodzi o niskie częstotliwości (podobnie jak u innych dużych zwierząt).
Jak u innych celurozaurów, dość dobrze rozwinięty był zmysł równowagi, pozwalający na aktywne życie - szybkość, zwrotność i zwinność.
Jak wskazuje badanie Carbone'a i in. (2011), T. rex nie mógłby być wyłącznie padlinożercą, bo nie mógłby znaleźć wystarczającej ilości pożywienia - znalezienie małego zdechłego dinozaura (klasa wielkości ok. 75 kg) zajęłoby mu średnio ok. 6 dni, dużego (ok. 700 kg) - 55 dni, a pięciotonowego ponad rok! Oczywiście to bardzo niepewne szacunki, oparte na wielu uproszczeniach i niewiadomych, ale obrazują skalę problemu, jaki miałby padlinożerny T. rex przy żywieniu się takim pożywieniem.
W ekosystemie musi być jakiś duży, czołowy drapieżnik. Jeśli tyranozaurydy były padlinożerne, to w ich sąsiedztwie musiały żyć jakieś duże, nieznane superdrapieżniki. Ekosystem tak produktywny jak Serengeti dostarczałby wystarczającej ilości pożywienia padlinożernym tyranozaurydom, ale tylko gdyby były one zmiennocieplne. Sugeruje się, że współczesne ekosystemy nie posiadają wielkich lądowych padlinożerców, gdyż ptaki padlinożerne o wiele lepiej radzą sobie ze zdobywaniem padliny. Niektórzy podejrzewają, że duże pterozaury były padlinożercami. Ich ogromne skrzydła pozwalały im szybować bez żadnego wysiłku, wyglądając padliny. W takim przypadku dla tyranozaurydów brakowałoby pokarmu. Na znalezionej w Montanie miednicy triceratopsa znaleziono ponad 50 śladów ugryzień. Wykonanie odlewów wykazało, że pozostawił je tyranozaur. Nie wiadomo jednak czy zabił on owego triceratopsa, czy po prostu pożywiał się jego padliną. Lambe w 1917 roku opisał dobrze zachowany szkielet gorgozaura. Sugerując się brakiem zużyć na zębach, stwierdził, że był on padlinożercą. Jednak dzisiaj wiemy, że teropody szybko wymieniały zęby, tak jak rekiny i krokodyle. Młode tyranozaurydy miały bardzo długie nogi. Mogły one zaganiać zdobycz w kierunku dorosłych, które zabijały ją ugryzieniem szczęk. Dorosły tyranozaur mógł biegać z prędkością ok. 40 km/h. Sugeruje się również, że mógł on osiągać prędkość jedynie 18 km/h albo nawet ponad 70 km/h. Jednak jak później wykazano, ta ostatnia opcja jest mało prawdopodobna, gdyż aby przykładowy tyranozaur mógł biec z taką prędkością, potrzebowałby mięśni nóg o łącznej masie wynoszącej ok. 40% całkowitej masy ciała. Tak więc mógł biegać szybciej niż większość ludzi. Jak na zwierzę o wadze sześciu ton jest to duża prędkość. Równocześnie potężna czaszka tyranozaura nie była w pełni równoważona, co ograniczało jego zwinność - wg autorów filmu "Dinozaury zabójcy" był jeszcze mniej zwinny niż Triceratops. Jednak wraz ze wzrostem prędkości biegu rośnie ryzyko wywrócenia się zwierzęcia. Tyranozaurydy miały zredukowane kończyny przednie, więc nie jest wyjaśnione, jak po upadku mogły się podnieść. Równocześnie sugeruje się, że tyranozaurydy były najszybszymi z wielkich teropodów. Jack Horner i Don Lessem (1993) stwierdzili, że tyranozaur był powolny i niezdolny do biegu, gdyż jego stosunek długości kości udowej do piszczeli wynosi ~1. U dinozaurów, u których można przypuszczać, że były szybkie np. u troodonta było odwrotnie. John Ostrom - znany z ponownego przytoczenia teorii dinozaurowego pochodzenia ptaków (którą naprawdę sformułował Thomas Henry Huxley, jednak została ona odrzucona przez ówczesne środowisko naukowe) dowiódł, że stosunek kości udowej do piszczelowej nie jest tak ważny przy ustalaniu prędkości biegu, jak stosunek stopy do kości piszczelowej. U współczesnych szybko biegających ptaków wynosi on 0,85, u Sinornithomimus - 0,88, a u Deinonychus - 0,48. Z kolei u największych (11-13 m) tyranozaurów jest to 0,51-0,59, co jest dobrym wynikiem na tle teropodów nawet mniejszych teropodów (proporcja ta zmniejsza się wraz ze wzrostem): Acrocanthosaurus (10 m) - 0,43-0,46, Chilantaisaurus (11 m) - 0,43-0,48 i Allosaurus (9,5 m) - 0,52. U żyrafy biegnącej ok. 50 km/h w bezpiecznym środowisku np. w zoo szansa na złamanie nogi lub inny uraz jest duża, a u żyjącego dziko tyranozaura i innych tyranozaurydów powinna być jeszcze większa. Tułów tyranozaura znajdował się ok. 1,5 m nad ziemią, przy przewróceniu się tyranozaur uderzyłby w ziemię z przeciążeniem równym 6 G (sześciokrotnie większym od przyśpieszania ziemskiego). Niewielkie przednie łapy nie mogły amortyzować uderzenia. Równocześnie w pobliżu tyranozaura i innych tyranozaurydów żyły powolne dinozaury, takie jak ankylozaur czy triceratops, jeszcze wolniejsze od tyranozaura. Mógł on też polować z zasadzki na szybsze zwierzęta np. hadrozaurydy.
Jeden ze szkieletów edmontozaura ma uszkodzone przez ugryzienie tyranozaura kręgi ogonowe. Gojenie się ich wskazuje, że zostały one zadane za życia edmontozaura - czyli tyranozaur próbował go upolować. Na jednej z czaszek triceratopsa widać charakterystyczne ślady powstałe w wyniku ugryzienia tyranozaura. Zagęszczenie tkanki świadczy o gojeniu się rany, z czego jasno wynika, że tyranozaur walczył z tym triceratopsem.
Jak u wielu innych mięsożerców, także wśród nieptasich teropodów mogło dochodzić do kanibalizmu, lecz dotąd nie wykazano tego z całą pewnością (w przypadku Majungasaurus zob. artykuł o Abelisauridae - wymiary). Longrich i in. (2010) donieśli aż o czterech kościach ze śladami ugryzień przypisanych do T. rex - co stanowi aż ¼ wszystkich, które noszą takie wyżłobienia. Trzy z nich to kości stopy, czwarta to kość ramienna - co świadczy o tym, że doszło do odżywania się tyranozaurzą padliną. Nie ma jednak zupełnej pewności, czy był to kanibalizm - w tym czasie na tym terenie żył też inny, lecz znacznie rzadszy tyranozauryd (nanotyran).
Podsumowując - tyranozauroidy były przystosowanymi do polowania na duże dinozaury drapieżnikami. Widać wśród nich specjalizację - wczesne (guanlong, stokesozaur) oraz nieduże i lekko zbudowane (alioram, ksiongguanlong) oraz młode polowały zapewne na mniejsze zwierzęta, a duże, masywne i dorosłe (tyranozaur, daspletozaur) potrafiły upolować dużą zwierzynę, miażdżąc jej kości. Wielkie tyranozaury, znane z możliwości do przegryzania i zjadania kości, były też zdolne do delikatnego obierania kości z mięsa (tarbozaur - Hone i Watabe, 2010).
Zachowania społeczne
Na pysku młodego daspletozaura oznaczonego TMP 94.143.1 widać ślady ugryzień innego przedstawiciela tego rodzaju. Podobne uszkodzenia widać na pysku innego okazu, ale tym razem należącego do dorosłego daspletozaura. Ich zagojenie świadczy, że zostały zadane jeszcze za życia zwierzęcia. W formacji Two Medicine znaleziono trzy szkielety daspletozaurów, w tym dwóch średnich rozmiarów i jednego dorosłego osobnika. Wokół nich odkryto szczątki pięciu hadrozaurydów. Wszystko wskazuje, że ciała wymienionych wyżej gadów nie zostały zniesione w jedno miejsce przez jakiś kataklizm, jak np. powódź, ale przysypane osadami tym samym czasie i miejscu. Przyczyna ich śmierci pozostaje tajemnicą. W cmentarzysku Dry Island znaleziono szczątki co najmniej dwunastu albertozaurów w różnym wieku. Ponieważ wokół nie znaleziono szczątków żadnych roślinożerców, wątpliwe jest, aby w jednym miejscu znalazły się niezależnie zwabione zapachem padliny. Neguje się jednak to, że wymienione wyżej znaleziska świadczą o opiece rodzicielskiej i życiu w grupach u tyranozaurydów. W 1997 r. odkryto kompletny szkielet triceratopsa znany jako "Kelsey". Wokół niego znajduje się ponad dwadzieścia zębów pochodzących od tyranozaura bądź nanotyrana (synonimów?). Niezbyt prawdopodobne wydaje się, aby jeden tyranozaur/nanotyran stracił tyle zębów podczas jednego posiłku. Stanowi więc to kolejną przesłankę za życiem w stadach tyranozaurydów. Wokół samicy tyranozaura nazwanej "Sue" znajdują się szczątki osobników młodych. Natomiast na kręgach szyjnych innego tyranozaura - "Stana" widać ślady ugryzień innego "jaszczurzego tyrana". Więcej na ten temat w osobnym artykule poświęconym zachowaniu teropodów, a będącym obecnie w przygotowaniu.
Rozwój osobniczy
Gregory Erickson i jego współpracownicy przebadali rozwój i przebieg życia tyranozaurydów. Tempo wzrostu może być badane za pomocą oględzin tzw. linii wzrostu wewnątrz kości, analogicznych z pierścieniami występującymi w pniach drzew. Ich badanie wykazało, że młode tyranozaurydy przechodziły fazę bardzo szybkiego wzrostu mniej więcej w środku życia, przez około cztery lata. Wtedy to szczęki nabierały ostatecznego kształtu (tj. stawały się wysokie i szerokie), kończyny tylne proporcjonalnie się skracały, całe ciało nabierało masywniejszego budowy i przypuszczalnie dochodziło do utraty piór. Po osiągnięciu dojrzałości płciowej tempo wzrostu tyranozaurydów znacząco malało, jednak rosły one do końca życia. Najmniejszy znany okaz tyranozaura (LACM 28471 znany jako "Jordan" miał ok. 30 kg wagi i był w chwili śmierci w wieku dwóch lat. Tymczasem największy znany dość kompletny okaz tyranozaura (FMNH PR 2081, "Sue") ważył najprawdopodobniej ponad 5400 kg. Młode tyranozaury do wieku 14 lat nie przekraczały ok. 1,8 t masy, ale podczas fazy szybkiego rozwoju rocznie przybierały ok. 600 kg, co daje mniej więcej 2 kilogramy dziennie. Wiek "Sue" jest szacowany na 28 lat. Jednak czy jest ona faktycznie największym okazem tyranozaura, a tym samym tyranozauroida? Jack Horner i jego zespół w 2000 odkrył szkielet tyranozaura nazwany "Celeste" lub "C-rex". Szacuje się, że jest on 10% razy większy od Sue. Inne tyranozaurydy nie przybierały na wadze aż tak szybko. Dla porównania daspletozaur w czasie okresu szybkiego dorastania mógł osiągać rocznie maksymalny wzrost masy 180 kg. Masę tę obliczono, posługując się oszacowaniem masy dorosłego daspletozaura, wynoszącej 1800 kg. Jeśli jednak jego masa była inna (podawana jest też na 3600 kg lub 2500 kg) przyrost roczny był większy. U gorgozaura maksymalny przyrost w ciągu roku podczas tej fazy wynosił 110 kg. Próbujące samodzielnie polować na być może mniejsze ofiary młode tyranozaurydy zajmowały przypuszczalnie inne nisze ekologiczne niż dorosłe (tak jak u współczesnych waranów z Komodo, gdzie osobniki młode żyją na drzewach i polują na owady, podczas gdy dorosłe są naziemnymi drapieżnikami i padlinożercami). Może to tłumaczyć brak średniej wielkości drapieżników w późnokredowej Ameryce Północnej.
Jak już zostało wspomniane, tyranozaurydy po osiągnięciu dojrzałości płciowej kończyły fazę szybkiego wzrostu i rosły aż do śmierci, choć już bardzo powoli. Jednoczesne dojrzewanie płciowe i szybki wzrost wydaje się cechą powszechną u małych i dużych dinozaurów. Podobnie jest u współczesnych dużych ssaków, takich jak człowiek czy słonie. Natomiast u ptaków - najbliższych współczesnych krewnych tyranozaurydów - występuję odwrotna sytuacja - dojrzałość zaczyna być osiągana po osiągnięciu pełnych rozmiarów. Erickson razem ze współpracownikami porównali ilość okazów tyranozaurydów w danym wieku. Znaleziono niewiele tyranozaurydów będących przed fazą szybkiego wzrostu, natomiast dość dużo szczątków tyranozaurydów w tym okresie i dorosłych. Może to sugerować niewielką śmiertelność młodych, ale ponad dwuletnich, tyranozaurydów (wśród młodszych śmiertelność była prawdopodobnie bardzo wysoka). Współcześnie spotykana jest ona u większych ssaków, jak np. słonie. U przedstawicieli Tyrannosauridae mogła wynikać ona z nieobecności dużych nie-tyranozaurydowych drapieżników w ekosystemie lub ochrony rodzicielskiej. Niektórzy spekulują, że może to świadczyć o życiu w grupkach (rodzinnych?) tyranozaurydów, gdyż ich zdaniem w skupiskach zawsze zostawała wystarczającą ilość pożywienia dla osobników młodych. Należy jednak pamiętać, że rzadkość znalezisk młodych tyranozaurydów wcale nie musi świadczyć o ich niskiej śmiertelności - mogły się one po prostu nie zachować w zapisie kopalnym. Wiele tyranozaurydów ginęło krótko (ok. sześciu lat) po osiągnięciu dojrzałości płciowej, tak jak u niektórych dzisiejszych długowiecznych ptaków i ssaków, których współczesne gatunki charakteryzują się niską śmiertelnością młodych i wysoką osobników wkrótce po fazie dojrzewania płciowego.
Wymiary:
>0,75-1 m (Aviatyrannis), >1,6 m (Dilong), ?~2,4 m (?Proceratosaurus), 2-3 m (Stokesosaurus clevelandi, >2,5 m (Raptorex) <<< ~13 m (Tyrannosaurus)
Jurajskie tyranozauroidy były niewielkie - mierzyły przeważnie 2-3 m (Stokesosaurus clevelandi, Guanlong, Proceratosaurus - o ile jest tyranozauroidem), lecz niektóre były większe (ponad 4 m długości u Kileskus i 4-6 m u Stokesosaurus langhami). Co ciekawe, w ewolucji jurajskich proceratozaurydów (o ile jest to grupa monofiletyczna) można dostrzec miniaturyzację - bardziej bazalne są większe. Maleńki był Aviatyrannis, lecz całkiem możliwe, że odnaleziony materiał pochodził od młodych osobników (obliczenia oparto na wymiarach najbardziej kompletnej kości biodrowej, inne były trochę większe). Może był wtórnie zminiaturyzowany tak jak Dilong (>1,6 m), którego odnalezione osobniki nie były w pełni wyrośnięte. Większość wczesnokredowych tyranozauroidów (Xiongguanlong i Eotyrannus) osiągało podobne rozmiary co ich najwięksi jurajscy krewni. Najbardziej zróżnicowane pod względem rozmiarów były one właśnie wtedy. Sinotyrannus miał zdaniem Li i in. (2009) aż 9-10 m długości, metrową czaszkę i 77-centymetrową kość biodrową, co czyni go największym znanym wczesnokredowym tyranozauroidem, a zarazem teropodem z grupy Jehol. Paul (2010) nieoficjalnie ocenia go na 9 m. Szacunki na podstawie wymiarów krewniaków wskazują jednak na mniejsze rozmiary - ok. 8-9 m (6,5-8 m dla Guanlong, 9-9,5 m dla Dilong lub 7-9 m dla Albertosaurus, Gorgosaurus i Daspletosaurus, przy czym górna granica dotyczący porównań dla czaszki a dolna dla kości biodrowej - dużo krótszej w porównaniu do czaszki niż u innych tyranozauroidów; szacunki dla mniejszych krewniaków mogą być przeszacowane, ponieważ duże teropody mają proporcjonalnie większe czaszki. Natomiast porównania dla tyranozaurydów mogą być niedoszacowane, gdyż ich czaszki były proporcjonalnie większe niż u bardziej bazalnych tyranozauroidów, co których należy Sinotyrannus). Cau (online) przedstawił wyniki swojej nieoficjalnej analizy kladystycznej, wg której jest on prymitywnymi tyranozauroidem, blisko spokrewnionym ze stokesozaurem. Może to świadczyć, że ten klad (opierający się wszakże jedynie na jednej synapomorfii) w ciągu swej ewolucji dążył do powiększenia rozmiarów.
Większość tyranozaurydów (i Bistahieversor, który może być jednym z nich) osiągała podobne rozmiary co sinotyran, lecz niektóre były jeszcze większe (Tarbosaurus, Zhuchengtyrannus i szczególnie Tyrannosaurus). Proces ten doszedł do szczytu w Ameryce Północnej.
Gdzie:
Laurazja (Ameryka Północna, Azja, Europa), Australia, ?Madagskar
Jak inne grupy celurozaurów, także tyranozauroidy wywodzą się prawdopodobnie z Azji, skąd znana jest też większość proceratozaurydów. Początkowo - w jurze i początku wczesnej kredy występowały prawdopodobnie w całej Laurazji. Zęby podobne do tyranozauroidowych znane są ze środkowej jury (baton, 168-164 Ma) Madagskaru (Maganuco i in., 2005). Kość łonowa należąca do - jak się wydaje - bardziej zaawansowanego od Raptorex i osiągającego ok. 3 m tyranozauroida pochodzi ze "środkowej" kredy (apt-alb, ~112 Ma) Australii (Benson i in., 2010 A; okaz NMV P186046). Przynależność tej skamieniałości do tyranozauroidów została zakwestionowana (Herne i in., 2010), lecz duża (>0.45 długości trzonu) dystalna ekspansja kości łonowej, obecna u wielu tetanurów w połączeniu z charakterystyczną dla celurozaurów wąskością tej kości wskazuje, że to rzeczywiście przedstawiciel omawianego kladu. Nie są znane żadne "środkowo-" i późnokredowe tyranozauroidy z Europy.
Tyranozaurydy pochodziły prawdopodobnie z zachodniej części z Ameryki Północnej, skąd zawędrowały do Azji i być może także wschodniej części Ameryki, odizolowanej wewnętrznym morzem (Appalachiosaurus, mogący być albertozaurynem).
Tyranozauroidy preferowały prawdopodobnie wilgotniejsze środowiska, podobnie jak niektóre inne grupy (Therizinosauria, Ornithomimosauria, Hadrosauroidea).
Kiedy:
~ 166 Ma -> 65 Ma
Najstarsze:
Kileskus 168-164 Ma
Najmłodsze:
Tyrannosaurus 68-65 Ma
Dowiedz się więcej:
- o bazalnych tyranozauroidach:
Aviatyrannis, ??Calamosaurus, ?Dilong, Dryptosaurus, ?Eotyrannus, "Futabasaurus", Raptorex, Stokesosaurus, Xiongguanlong
- ? w tym proceratozaurydach:
?Guanlong, Kileskus, Proceratosaurus, Sinotyrannus
- tyranozaurydach:
Alectrosaurus, (Aublysodon), (Deinodon)
- w tym albertozaurynach:
Albertosaurus, ??Appalachiosaurus, ??Bistahieversor, Gorgosaurus
- i tyranozaurynach:
?Alioramus, Daspletosaurus, Nanotyrannus, Tarbosaurus, Teratophoneus, Tyrannosaurus, Zhuchengtyrannus

Rekonstrukcja tyranozaura stworzona na potrzeby "Doliny tyranozaura". Obraz pochodzi z tej strony.

Wykres krzywych wzrostu tyranozaurydów. Zmodyfikowano z Erickson in. (2004). Pierwotny obraz pochodzi z tej strony

Rekonstrukcje czaszek tyranozauroidów. A - bazalny tyranozauroid Guanlong wucaii (autor: R. S. Li; z Xu i in., 2004), B - bazalny tyranozauroid Dilong paradoxus - IVPPV14243 (autorzy: R. S. Li, J. L. Huang i M.W. Yang; z Xu i in., 2004), C - zaawansowany tyranozauroid Raptorex kriegsteini - LH PV18 (autor: C. Abraczinskas; z Sereno i in., 2009), D - zaawansowany tyranozauroid lub bazalny tyranozauryn Alioramus altai - IGM 100/1844 (autor: Frank Ippolito; z Brusatte i in., 2009), E - albertozauryn Gorgorsaurus libratus - kompilacja niedorosłych AMNH 5664, CMN 11315, ROM 1247, TMP 91.36.500 I USNM 12815 (autor: Dino Pulerà, rysunek pochodzi z tej strony; z Carr, 1999), F - tyranozauryn Tarbosaurus bataar (autor: Karol Sabath; z Hurum i Sabath, 2003).

Rekonstrukcje szkieletów tyranozauroidów. Od góry: bazalny tyranozauroid Dilong paradoxus (autor: Ville Sinkkonen, rysunek pochodzi z tej strony), zaawansowany tyranozauroid Raptorex kriegsteini - młody okaz (z Sereno i in., 2009), albertozauryn Gorgorsaurus libratus - okaz młodociany (autor: Scott Hartman, rysunek pochodzi z tej strony) i zaawansowany tyranozauryd Tyrannosaurus rex (autor: Scott Hartman, rysunek pochodzi z tej strony).
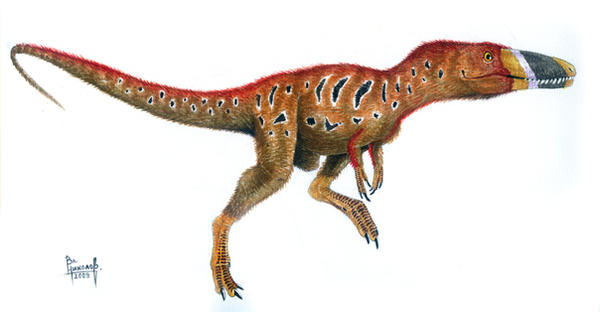
Rekonstrukcja Xiongguanlong. Autor: Vladimir Nikolov. Obrazek pochodzi ztej strony.

Kladogram z Currie i in., 2003 (tylko cechy czaszki).

Skalibrowany stratygraficznie kladogram z Holtz, 2004 (=Holtz i in., 2004; zob. art. o Tetanurae). Przerywaną linią ukazano dwie alternatywne pozycje Alioramus. Skala czasu za Walker i Geissman, 2009.

Skalibrowany stratygraficznie kladogram z Benson, 2008 (na podstawie Xu i in., 2006, z dodanymi Stokesosaurus, Eotyrannus i Deltadromeus). Skala czasu za Walker i Geissman, 2009.

Skalibrowany stratygraficznie kladogram z Sereno i in., 2009. Skala czasu za Walker i Geissman, 2009.

Skalibrowany stratygraficznie kladogram z Carr i Williamson, 2010 ukazujący też synapomorfie wybranych kladów. Cf. Alectrosaurus to nienazwany tyranozauroid (GIN 100/50, 100) a BYU 8120 i in. to Teratophoneus. Alioramus zawiera tylko A. remotus a Stokesosaurus - S. clevelandi. Skala czasu za Ogg i in., 2004.

Skalibrowany stratygraficznie kladogram z Brusatte i in., 2009 (na podstawie Carr i Williamson, 2010 z dodanym Alioramus altai i Xiongguanlong). Xiongguanlong następnie usunięto, gdyż tworzył politomię z Eotyrannus, Appalachiosaurus, Dryptosaurus i Albertosaurinae+Tyrannosaurinae. "Takson z Nowego Meksyku" (obecnie Bistahieversor) był wtedy siostrzanym taksonem Albertosaurus. "Takson z Utah" to Teratophoneus.
Skala czasu za Walker i Geissman, 2009.

Skalibrowany stratygraficznie kladogram z Brusatte i in., 2009 (z Xiongguanlong i bez Allosaurus - Cau, online). "Takson z Nowego Meksyku" to Bistahieversor. "Takson z Utah" to Teratophoneus. Skala czasu za Walker i Geissman, 2009.

Kladogram z Averianov i in., 2010 (na podstawie Smith i in., 2008 z dodanymi Proceratosaurus, Kileskus i Guanlong i odjętymi Tugulusaurus i NMV P186076. Usunięto też 6 nieinformatywnych cech).

Skalibrowany stratygraficznie kladogram z Brusatte i in., 2010B na podstawie analizy Brusattego i Carra. "Utah taxon" to Teratophoneus.

Skalibrowany stratygraficznie kladogram z Carr i in., 2011 ukazujący też synapomorfie wybranych kladów (na podstawie Brusatte i in., 2009).

Kladogram z Zanno i Makovicky, 2011 (na podstawie Sereno i in., 2009 z dodanym FMNH PR 2750 i 4 nowymi cechami).
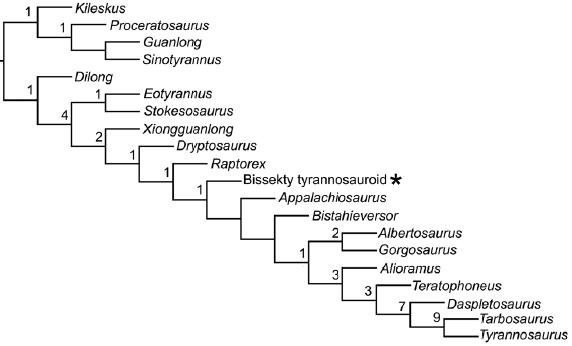
Kladogram z Averianov i Sues, 2012 (na podstawie Brusatte i in., 2009 z dodanym materiałem z turonu Uzbekistanu).

Skalibrowany stratygraficznie i względem rozmiarów kladogram z Cau (online, na podstawie matrycy Cau i Maganuco, w przygotowaniu), wskazujący na czterokrotne, niezależne powiększanie rozmiarów tyranozauroidów. Z lewej czas (w Ma), na dole długość dorosłych osobników. Obrazek pochodzi z tej strony (kliknij na niego aby powiększyć).

Kladogram z Carr, 2005 (tylko cechy kończyn tylnych).
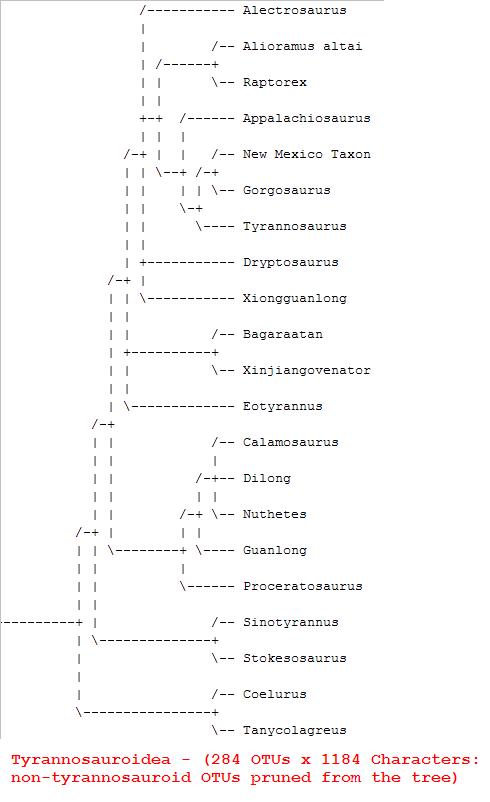
Kladogram z Cau (online).
Źródła:
Publikacje naukowe:
Averianov, A.O., Krasnolutskii, S.A. & Ivantsov, S.V. (2010) "A new basal coelurosaur (Dinosauria: Theropoda) from the Middle Jurassic of Siberia" Proceedings of the Zoological Institute RAS 314, 42-57.
Averianov, A. & Sues, H.-D. (2012) "Skeletal remains of Tyrannosauroidea (Dinosauria: Theropoda) from the Bissekty Formation (Upper Cretaceous: Turonian) of Uzbekistan" Cretaceous Research, doi:10.1016/j.cretres.2011.11.009
Benson, R.B.J. (2008) "New information on Stokesosaurus, a tyrannosauroid (Dinosauria: Theropoda) from North America and the United Kingdom" Journal of Vertebrate Paleontology, 28, 732-750.
Benson, R.B.J., Barrett, P.M., Rich, T.H. & Vickers-Rich, P. (2010 A) "A Southern Tyrant Reptile" Science, 327, 1613.
Benson, R.B.J., Barrett, P.M., Rich, T.H., Vickers-Rich, P., Pickering, D. & Holland, T. (2010 B) "Response to Comment on "A Southern Tyrant Reptile"" Science, 329, 1013-d
Brusatte, S.L., Carr, T.D., Erickson, G.M., Bever, G.S. & Norell, M.A. (2009) "A long-snouted, multihorned tyrannosaurid from the Late Cretaceous of Mongolia" Proceedings of the National Academy of Sciences, 106, 17261-17266.
Brusatte, S.L., Chure, D.J., Benson, R.B.J. & Xu, X. (2010A) "The osteology of Shaochilong maortuensis, a carcharodontosaurid (Dinosauria: Theropoda) from the Late Cretaceous of Asia" Zootaxa, 2334, 1-46.
Brusatte, S.L., Norell, M.A., Carr, T.D., Erickson, G.M., Hutchinson, J.R., Balanoff, A.M., Bever, G.S., Choiniere, J.N., Makovicky, P.J. & Xu, X. (2010B) "Review: Tyrannosaur Paleobiology: New Research on Ancient Exemplar Organisms" Science, 329, 1481-1485.
Brusatte, S.L., Benson, R.B.J. & Norell, M.A. (2011) "The Anatomy of Dryptosaurus aquilunguis (Dinosauria: Theropoda) and a Review of Its Tyrannosauroid Affinities" American Museum Novitates, 3717, 1-53. doi: 10.1206/3717.2
Carbone, C., Turvey, S.T. & Bielby, J. (2011) "Intra-guild competition and its implications for one of the biggest terrestrial predators, Tyrannosaurus rex" Proceedings of the Royal Society of London B: Biological Sciences, 278, 2682-2690. doi: 10.1098/rspb.2010.2497
Carr, T.D., Williamson, T.E., Britt, B.B. & Stadtman, K. (2011) "Evidence for high taxonomic and morphologic tyrannosauroid diversity in the Late Cretaceous (Late Campanian) of the American Southwest and a new short-skulled tyrannosaurid from the Kaiparowits formation of Utah" Naturwissenschaften, 98(3), 241-246. doi: 10.1007/s00114-011-0762-7
Carr, T.D. & Williamson, T.E. (2010) "Bistahieversor sealeyi, gen. et sp. nov., a new tyrannosauroid from New Mexico and the origin of deep snouts in Tyrannosauroidea" Journal of Vertebrate Paleontology, 30, 1-16.
Choiniere, J.N., Clark, J.M., Forster, C.A. & Xu, X. (2010) "A basal coelurosaur (Dinosauria: Theropoda) from the Late Jurassic (Oxfordian) of the Shishugou Formation in Wucaiwan, People's Republic of China" Journal of Vertebrate Paleontology, 30(6), 1773-1796. doi: 10.1080/02724634.2010.520779
Csiki, Z., Vremir, M., Brusatte, S.L. & Norell, M.A. (2010) "An aberrant island-dwelling theropod dinosaur from the Late Cretaceous of Romania" Proceedings of the National Academy of Sciences, 107(35), 15357-15361, doi: 10.1073/pnas.1006970107
Fowler, D.W., Woodward, H.N., Freedman, E.A., Larson, P.L. & Horner, J.R. (2011) "Reanalysis of “Raptorex kriegsteini”: A Juvenile Tyrannosaurid Dinosaur from Mongolia" PLoS ONE 6(6): e21376. doi:10.1371/journal.pone.0021376
Herne, M.C., Nair, J.P & Salisbury, S.W. (2010) "Comment on "A Southern Tyrant Reptile"" Science, 329, 1013-c.
Holtz, T.R. (1994) "The phylogenetic position of the Tyrannosauridae: implications for theropod systematics" Journal of Paleontology, 68, 1100-1117.
Holtz, T.R. (2004) "Tyrannosauroidea" [w:] Weishampel, D.B., Dodson, P. & Osmólska, H. "The Dinosauria" Berkeley and Los Angeles: University of California Press, 111-136.
Hone, D.W.E. & Watabe, M. (2010) "New information on scavenging and selective feeding behaviour of tyrannosaurids" Acta Palaeontologica Polonica, 55 (4), 627-634. doi:10.4202/app.2009.0133
Ji, Q., Ji, S.A. & Zhang, L.-J. (2009) "First large tyrannosauroid theropod from the Early Cretaceous Jehol Biota in northeastern China" Geological Bulletin of China, 28, 1369-1374.
Lee, M.S.Y. & Worthy, T.H. (2011) "Likelihood reinstates Archaeopteryx as a primitive bird" Biology Letters. doi: 10.1098/rsbl.2011.0884
Longrich, N.R., Horner, J.R., Erickson, G.M., Currie, P.J. (2010) "Cannibalism in Tyrannosaurus rex" PLoS ONE 5(10): e13419. doi:10.1371/journal.pone.0013419
Maganuco, S., Cau A. & Pasini G. (2005) "First description of theropod remains from the Middle Jurassic (Bathonian) of Madagascar" Atti della Societa Italiana di Scienze Naturali e del Museo Civico di Storia Naturale in Milano, 146, 165-202.
Makovicky, P.J., Li, D., Gao, K-Q., Lewin, M., Erickson G.M. & Norell, M.A. (2010) "A giant ornithomimosaur from the Early Cretaceous of China" Proceedings of the Royal Society B: Biological Sciences, 277, 191-198. doi: 10.1098/rspb.2009.0236
Naish, D. & Martill, D.M. (2007) "Dinosaurs of Great Britain and the role of the Geological Society of London in their discovery: basal Dinosauria and Saurischia" Journal of the Geological Society, 164, 93-510.
Perez-Moreno, B.P., Sanz, J.L., Buscalioni, A.D., Moratalla, J.J., Ortega, F. & Raskin-Gutman, D. (1994) "A unique multitoothed ornithomimosaur from the Lower Cretaceous of Spain" Nature, 370, 363-367.
Rauhut, O.W.M. (2003). "A tyrannosauroid dinosaur from the Upper Jurassic of Portugal" Palaeontology, 46, 903-910.
Rauhut, O.W.M., Milner, A.C. & Moore-Fay, S. (2010) "Cranial osteology and phylogenetic position of the theropod dinosaur Proceratosaurus bradleyi (Woodward, 1910) from the Middle Jurassic of England" Zoological Journal of the Linnean Society, 158, 155-195.
Senter, P. (2010) "Using creation science to demonstrate evolution: application of a creationist method for visualizing gaps in the fossil record to a phylogenetic study of coelurosaurian dinosaurs" Journal of Evolutionary Biology, 23, 1732-1743.
Sereno, P.C., Tan, L., Brusatte, S.L., Kriegstein, H.J., Zhao, X. & Cloward, C. (2009) "Tyrannosaurid Skeletal Design First Evolved at Small Body Size" Science, 326, 418-422.
Stevens, K.A. (2006) "Binocular vision in theropod dinosaurs" Journal of Vertebrate Paleontology, 26 (2), 321-330.
Turner, A.H., Pol, D., Clarke, J.A., Erickson, G.M., & Norell, M. (2007) "A basal dromaeosaurid and size evolution preceding avian flight" Science 317, 1378-1381.
Turner, A.H., Pol, D. & Norell, M.A. (2011) "Anatomy of Mahakala omnogovae (Theropoda: Dromaeosauridae) Tögrögiin Shiree, Mongolia" American Museum Novitates, 3722, 1-66. doi: 10.1206/3722.2
Witmer, L.M. & Ridgely, R.C. (2009) "New Insights Into the Brain, Braincase, and Ear Region of Tyrannosaurs (Dinosauria, Theropoda), with Implications for Sensory Organization and Behavior" The Anatomical Record: Advances in Integrative Anatomy and Evolutionary Biology, 292, 1266-1296. doi: 10.1002/ar.20983
Xu, X., Zheng, X. & You, H. (2010) "Exceptional dinosaur fossils show ontogenetic development of early feathers" Nature, 464, 1338-1341.
Zanno, L.E. & Makovicky, P.J. (2011) "On the earliest record of Cretaceous tyrannosauroids in western North America: implications for an Early Cretaceous Laurasian interchange event" Historical Biology, doi: 10.1080/08912963.2010.543952
Zelenitsky, D.K, Therrien, F. & Kobayashi, Y. (2009) "Olfactory acuity in theropods: palaeobiological and evolutionary implications" Proceedings of the Royal Society B, 276, 667-673. doi: 10.1098/rspb.2008.1075
Inne:
Carr, T.D. (2005) "Phylogeny of Tyrannosauroidea (Dinosauria: Coelurosauria) with special reference to North American forms" Dysertacja doktorska, University of Toronto
Miyashita, T. & Currie, P. (2009) "A new phylogeny of the Tyrannosauroidea (Dinosauria, Theropoda)" Journal of Vertebrate Paleontology, 29, 149A [abstrakt]
Paul, G.S. (2010) "The Princeton Field Guide to Dinosaurs" wyd. Princeton University
Williams, S., Brusatte, S., Mathews, J. & Currie, P. (2010) "A new juvenile Tyrannosaurus and a reassessment of ontogenetic and phylogenetic changes in tyrannosauroid forelimb proportions" Society of Vertebrate Paleontology Seventienth Anniversary Meeting Program and Abstracts Book, 188A [abstrakt]
http://dml.cmnh.org/1998Jun/msg00538.html
http://qilong.wordpress.com/2010/08/27/ ... -or-is-it/ (komentarz 1)
http://dml.cmnh.org/2011Oct/msg00220.html











 Paleontologia (i nauka ogólnie) tak nie działa. Timimus ma pewne cechy tyranozauroidów, ale jest za bardzo delikatny (wg niżej cytowanych autorów). Chodzi o celurozaura, ale dalej to już jest niepewne.
Paleontologia (i nauka ogólnie) tak nie działa. Timimus ma pewne cechy tyranozauroidów, ale jest za bardzo delikatny (wg niżej cytowanych autorów). Chodzi o celurozaura, ale dalej to już jest niepewne.{kind=link}
{kind=link}